Héjas gyümölcsfajok nemesítésének nemzetközi irányvonalai, fontosabb fajtaújdonságai
Szerző: Bujdosó Géza
Dió (Juglans regia L.)
A diónemesítés rövid története
A Juglans nemzettségbe 21 faj tartozik, melyek megtalálhatók a világ szinte minden pontján. E fajok közül legnagyobb jelentőségű a Juglans regia L., ezért kizárólag e fajjal foglalkozunk ebben a fejezetben.
A Juglans regia Közép-Ázsia hegyvidéki erdeiben honos, megtalálható a nyugat-kínai Xinjiang tartományban, Kazahsztán, Üzbegisztán és Dél-Kirgizisztán egyes területein, továbbá Nepál, Tibet, Észak-India és Nyugat-Pakisztán hegyvidéki területeitől Afganisztánon, Türkmenisztánon és Iránon keresztül egészen Azerbajdzsán, Örményország, Grúzia és Kelet-Törökország egyes régióiban is. Kelet-Európában és a Balkán félsziget országaiban is ősidők óta van néhány iráni, illetve kelet-török eredetű dió maradványpopuláció, melyek az ókori görög kereskedők és telepesek révén kerültek ide (Zohary és Hopf, 1993, Terpó 1976). Az ókori Rómában Jupiter magjának nevezték a diót, innen ered a Juglans nemzetség név. Az ókori rómaiak tovább terjesztették a diót Európában, Olaszország után Franciaországban, Spanyolországban, Portugáliában és Németország déli részén is ismerté vált ez a gyümölcsfaj (Leslie és McGranahan, 1998). A mai Amerikai Egyesült Államok területére az angol telepesek vitték az első diófákat 1562-ben, innen származik az „English walnut” azaz angol dió elnevezés, mivel Észak-Amerikában a Juglans nigra L. őshonos (McGranahan és Leslie 2009).
A nemesítés első lépésének a „népi szelekciót” tekintjük, mikor számos jó tulajdonsággal rendelkező genotípust kezdtek szaporítani – sajnálatos módon – generatív úton. Mivel a dió nem magnemes, így számos értékes genotípus elveszett a generatív szaporítás következtében. Messze földön ismertek voltak a Kárpát-medencében kiválasztott első „fajták”: a Sebeshelyi gömbölyű, a Sebeshelyi hosszú és a Milotai, melyek kiváló termésminőséggel rendelkeztek.
Az egykori Jugoszláviában egészen sajátosan alakult ki a diófajta-szortiment. A dió nemesítése 1962-ben kezdődött a Novi Sad-i Egyetemen, ahol főleg a Vojvodina tartományban található diófákat értékelték és gyűjtötték az értékes genotípusokat további vizsgálat céljából. Hamar kiderült, hogy szükség lenne egy szélesebb körű mintagyűjtésre, ezért 1973-ban versenyt hirdettek „Keressük a minőségi diókat” címmel. Ez a „népi szelekció” alapozta meg az egykori jugoszláv diófajtaválasztékot, mivel nemcsak Vojvodina tartományból, hanem az ország egész területéről küldték a mintákat (Korac et al 1986).
A tudatos magyar diónemesítés kezdetén, 1910-ben a legjelentősebb francia fajtákból nagy mennyiségű magot importált a Mezőgazdasági Minisztérium. A magoncokat az ország különböző termesztőtájaiba ültették ki, s a populáció értékelésére az 1950-es években került sor. Megállapítást nyert, hogy a „francia diórasszból származó populációk nem elég alkalmazkodóképesek a hazai kárpáti rassz termőhelyi adottságaihoz” (Szentiványi 1998). Ekkor ismerték fel a magyar diónemesítés fontosságát. A magyar diónemesítést id. Porpáczy Aladár indította a Fertődi Gyümölcstermesztési Kutató-Fejlesztő Intézet Nonprofit Közhasznú Kft. jogelődjében. E munka folytatódott 1950-től kezdve az Állami Gyümölcs- és Dísznövénytermesztési Kutató-Fejlesztő Közhasznú Nonprofit Kft. jogelődjeiben Érden (továbbiakban Érdi Kutató). Itt állította elő tájszelekcióval Szentiványi Péter az Alsószentiváni 117, a Milotai 10 és a Tiszacsécsi 83 államilag elismert fajtákat. Az ugyancsak tájszelekcióval előállított ’Tiszacsécsi 2’ később visszavonásra került.
A hazánktól keletebbre elhelyezkedő diótermesztő országokban (Ukrajna, Románia) nagyon népszerű a tájszelekció, mivel a kárpáti rasszból származó diópopulációk genetikai variabilitása igen nagy. Ebből adódóan szinte valamennyi államilag minősített diófajtát tájszelekcióval állítottak elő. Ezek a szelekciós helyre jellemző ökológiai adottságokra alkalmasak, melyek mellett biztonságosan és eredményesen termeszthetők. Ezt azért fontos tudni, mert a dió ökológiai adaptációs képessége minimális, azaz nehezen tolerálja a jelentősen eltérő ökológiai körülményeket (Szentiványi 2006).
Az 1970-es években a tájszelekció mellett a keresztezéses nemesítést is alkalmazták a magyar diónemesítési programban. Apai szülőként a Kaliforniai Davis Egyetemen nemesített Pedro fajtát vonták be a hibridizációba. Az érdi kutatóintézetben szinte valamennyi korszerű kaliforniai nemesítésű fajtát ’(’Tehama’, ’Serr’, ’Vina’) felhasználták a keresztezéses nemesítés során, de csak a ’Pedro’ örökítette legjobban a tulajdonságait, ezért kizárólag ezzel a fajtával folytatta a későbbi munkát Szentiványi Péter. A Milotai 10 és a Pedro fajták keresztezéséből állította elő Szentiványi Péter a Milotai bőtermő, Milotai kései és a Milotai intenzív fajtákat. Az ’Alsószentiváni 117’ és a ’Pedro’ keresztezéséből a Bonifác és az Alsószentiváni kései fajták jöttek létre. Nagy reménnyel használták a biotikus és abiotikus toleranciával rendelkező Tiszacsécsi 83 fajtát is a keresztezéses nemesítési programban, de a ’Tiszacsécsi 83’ x ’Pedro’ kombinációkból nem keletkezett ígéretes genotípus.
Az USA-ban, a kaliforniai Davis Egyetemen szinte kizárólag keresztezéses nemesítéssel foglalkoznak, melynek eredménye a ‘Chandler’, a ‘Howard’ és a ‘Sunland’, melyek a mediterrán klímájú országok vezető diófajtáivá váltak (Hendricks et al 1998).

Mesterségesen beporzott nővirág (Fotó: Bujdosó)
Mutációval még nem állítottak elő államilag elismert diófajtát, ami valószínűleg azzal áll összefüggésben, hogy a diónak kicsi a mutációs hajlama.
Biotechnológiai módszerrel, génátvitellel állítottak elő néhány genotípust a Davis Egyetemen, de termesztésük még nem engedélyezett. Elképzelésük szerint e módszerrel mérsékelhető a magbél allergéntartalma, módosítható a fakadási idő, a koronahabitus, javítható a télállóság valamint a stressztűrés a hideggel és a hőséggel szemben (Gauthier és Jacobs 2011, Vahdati et al. 2002). A tannin, a telítetlen zsírsavak és a flavonoidok bioszintézisét kódoló gének térképezése jelenleg is tart (McGranahan és Leslie 2009). Továbbá Bacilus thuringiesnsis-t használnak a diót károsító rovarkártevők (Leslie et al. 2001) valamint az Agrobacterium fajok elleni rezisztencia elérése érdekében (Escobar et al. 2002).
A honosítás lehetőségének megítélése nem egyszerű a diófajták esetében. Úgy tűnik, hogy csak ott sikeres a honosítás, ahol az adott fajta nemesítési helyével megegyezőek vagy közel azonosak az ökológiai adottságok. Ez a tény aláhúzza a külföldön nemesített diófajták hazai adaptációs vizsgálatának fontosságát termesztésbe vonásuk előtt. Dióoltvány hiány miatt a magyar termesztők külföldről vásároltak szaporítóanyagot. Legtöbbször az egykori Csehszlovákiában nemesített Jupiter fajtát vásárolják a termesztők, de hazánkban a termés magja sötétté vált. Feltehetően a szárazabb és melegebb nyári időjárással áll összefüggésben ez a jelenség. A kaliforniai ‘Hartley’ honosítása sem volt sikeres Magyarországon, mivel ez a magyar fajtákénál hosszabb tenyészidővel és magasabb hőmérsékleti igénnyel rendelkezik, ezért kisméretű, sovány, kevésbé esztétikus magbél fejlődik belőle (Bujdosó 2003). Hozzá kell tenni, hogy egyik fajta esetében sincs tapasztalat öntözött körülmények között. A kaliforniai nemesítésű ‘Pedro’ honosítása sikeresnek mondható, így megtalálható a Nemzeti Fajtajegyzékben. Tudni kell viszont róla, hogy fakadási ideje középkorai (április 15–20 közötti), ezért csak a késő tavaszi fagyoktól védett termőhelyeken javasolt termeszteni.
Világviszonylatban eredményes honosításra példaként szolgálnak a kaliforniai diófajták (‘Chandler’, ‘Howard’, ‘Serr’), melyeket az Amerikai Egyesült Államokon kívül mediterrán klímájú országokban, Spanyolországban, valamint Ausztrália és Új-Zéland ökológiailag alkalmas részein is sikeresen termesztenek.
A magyar diófajtákat is sikeresen honosítják Németország középső és északi részén. Figyelemre méltó, hogy a terméshéj itt vékonyabb, ezért kíméletesebb betakarítást és áruvákészítést igényelnek. Megjegyzendő még, hogy a magyar fajták a diót károsító kórokozókkal szemben fogékonyabbak Észak- és Közép-Németországban (Németország északi részén kizárólag a három alapfajta terjedt el), valamint Bulgáriában és Örményországban is kedvező véleményeket mondanak róluk.

Izolált termesztési rendszerben tartott GMO diógenotípusok a Davis-i Egyetemen (Fotó: Bujdosó)
Fő nemesítési célok
Korábban valamennyi diótermesztő országban a legfontosabb nemesítési cél a kései fakadási idő és az oldalrügyeken való terméshozás elérése volt. Napjainkban e két nemesítési cél mellé még a korai érési idő is felzárkózott. Annak ellenére, hogy az északi féltekén a dió fő értékesítési időszaka a karácsonyi ünnepek előtt van, újabban a nagy diótermesztő országokban (USA, Franciaország, Románia) a héjas dió és a dióbél esetében is a mielőbbi piacra vitelt tartják fontosnak.
Természetesen elengedhetetlen a jó termésminőség is. Jelenleg a 32 mm-es termésátmérő számít az I. osztály alsó határának. Elvárt még, hogy a diófajtának kerek vagy gömbölyded alakja legyen. A hosszúkás alakú genotípusok kevésbé preferáltak, mivel átmérőjük nem éri el 32 mm-t. Fontos, hogy a terméshéj sima és világos színű legyen, s világos színű vagy világos alapon sötétbarna vékony erezettel rendelkezzen a bél. Továbbá 40%-os bélarány, jó törhetőség és tisztíthatóság, mellék- illetve utóíztől mentes jó íz, megfelelően zárt varrat az ideális diófajta legfontosabb ismérvei. Mivel a föld déli féltekéjén is egyre nagyobb mennyiségben termesztenek diót, és az onnan származó termés március – áprilisban érkezik Európába, egyre jobban előtérbe kerül az, hogy a dióbél kedvező világos színét minél hosszabban megőrizze. Emellett fontos még a rendszeres és bő terméshozás, a jó szaporíthatóság valamint a középerős növekedési erély is.
A Davis Egyetemen, a világ legnagyobb keresztezéses nemesítési programján belül a következőképpen fogalmazták meg az ideális diófajta kritériumait 2009-ben (McGranahan és Leslie 2009):
- kései fakadási idő,
- korai termőrefordulás, negyedik nyaras korban már legalább 500 kg legyen a szárított héjas termésmennyiség hektáronként,
- a csúcsrügyeken való terméshozás mellett nagyfokú oldalrügyön termőképesség,
- hajlam az alternatív termésképződésre (az apomixis előnyös, a partenokarpiát nem preferáljuk, mivel az ilyen diótermések nem tartalmaznak magbelet),
- legalább 0,6 t/ha szárított héjas termésmennyiség elérése termőkorban,
- rezisztentcia/tolerancia a diót károsító főbb kórokozókkal szemben,
- a termésérés időszaka október eleje előtt fejeződjön be,
- a sima terméshéjfelület, jól törhető és tisztítható termés, a bélarány ne legyen magasabb 50%-nál, a termés érje el a 30 mm-es termésátmérőt, világos színű magbél, a magbél tömege ideális, ha 8 és 9 g közötti, magbél könnyen törjön „féldiók”-ra,
- középerős növekedési erély,
A diótermesztésben jelenleg igen nagy növényvédelmi problémák vannak, melyért elsősorban a Xanthomonas arboricola pv. juglandis, másodsorban a BAN (Brown Apical Nekrosis) a felelős. A világ számos nemesítő műhelyében foglalkoznak a kutatók ezzel a problémával, de sajnos jelenleg még nem állítottak elő a dió xantomónászos betegségével szemben toleráns vagy rezisztens fajtát. A kutatók szerint akkor lehet toleráns fajtát előállítani, ha a helyi populációból emelünk ki egy olyan genotípust, mely toleranciát/rezisztenciát mutat e kórokozóval szemben (Rovira et al. 2007, Özaktan et al. 2007, 2008; Lovera et al. 2008, Solar et al. 2008; Tsiantos et al. 2008, Ruiz-García et al. 2009, Solar et al. 2009, Frutos és López 2012). Említésre méltó, hogy a nemzetközi kutatási tendenciákkal egyidőben a BCE Gyümölcstermő Növények Tanszékén is folyik a magyar diófajtáknak és néhány Erdélyben szelektált diógenotípusnak Xanthomonas baktériummal szembeni vizsgálata (Theisz et al. 2007).
A nemesítés genetikai forrásai
A Juglans regia L. (2n=32) mellett az Amerikai Egyesült Államokban nagymértékben használják a Juglans nigra L. valamint a Juglans hindsii Rehd. fajokat a nemesítési programban (McKenna és Epstein 2003). Kínában a Juglans regia mellett a kiváló szárazságtűrő, téltűrő és télálló Juglans manshurica Maxim, Juglans cathayensis Dode., meleg és nedves klímát jól tűrő, de alacsony téltűrőképességgel rendelkező Juglans sigillata Dode. a legjelentősebb génforrás (Tian et al 2009).
A magyar diónemesítés szerencsés helyzetben van, mert a kárpáti rassz egy kis populációja az országban megtalálható. Ebben a populációban igen nagy genetikai változatosság figyelhető meg, s többek között ennek köszönhetők a magyar nemesítés kiemelkedő eredményei. Egyrészt a magyar diófajta szortiment jellemezhető a legkorábbi érési idővel a Föld északi féltekéjén, mintegy 10–14 nappal megelőzve más diótermesztő országokat. Számos külföldi nemesítő műhelyben is hasonló korai érési idő elérése a cél. Másrészt a magyar diófajták legnagyobb termésméretükkel tűnnek ki, s ezáltal jelenleg prémium áruértéket képviselnek.
Interspecifikus hibridek megjelenése és termesztése elsősorban Kalifornia államra jellemző, ahol a Juglans regia x Juglans hindsii hibridjeként állították elő a ’Paradox’ alanyt (McGranahan és Leslie 2009). McKenna és Epstein (2003) felhívja a figyelmet, hogy a ’Paradox’ megjelölés nem minden esetben azonos szülőpároktól származó populációt jelölt, ami azt jelenti, hogy a kereskedelemben kapható ’Paradox’ alany a Juglans hindsii és a Juglans major vagy a Juglans hindsii és a Juglans nigra fajhibridje.
Több szerző kísérleti eredménye alapján kijelentik, hogy a fajhibridek alanykénti használata esetében igen fontos a szülőfajok ismerete. A Juglans nigra felhasználásával előállított fajhibridek sokkal fogékonyabbak az agrobaktériumos gyökérgolyvára Kaliforniában, mint a Juglans regia, a Juglans hindsii és a Juglans major populációkból előállítottak. Továbbá a Juglans cinerea és a Juglans ailantifolia keresztezéséből származó ’butterjaps’ (syn.: ’buartnuts’) valamint a Juglans nigra és a Juglans ailantifolia hibridjeként előállított ‘Leslie Burt’ alanyfajta is fogékony a dió xantomónaszos betegségére Kaliforniában (McKenna és Epstein 2003, McGranahan és Leslie 2009).
A fajhibridek jelentősége nemesfajták esetében a Cherry Leafroll potyvírus (továbbiakban CLRV) elleni védekezésben fontos, mivel csak a Juglans hindsii tartalmaz egy olyan domináns gént, mely a vírusfertőzés hatására hiperszenzitív reakciót vált ki a dióban. A Juglans hindsii x Juglans regia hibrideket Juglans regia-val visszakeresztezve 1:1 arányban találtak hiperszenzitív, illetve toleráns genotípusokat az utódpopulációban amerikai kutatók. Azt is el kell mondani az így létrejött fajhibridekről, hogy hímsterilek, így jelenleg még nyitott kérdés, hogy a CLRV-sal szemben hiperszenzitivitást mutató genotípusok hogyan hoznak termést, illetve a hiperszenzitív reakció működik-e a szabadföldön (McGranahan et al. 1997).
Kínában a legismertebb fajhibrid a Hebei tartományban lévő Taihang hegységben található Juglans hopeiensis Hu, mely a J. regia x J. mandshurica természetes hibridje. Ez a faj nemcsak gyümölcstermesztési jelentőséggel rendelkezik (termésmérete nagy, tetszetős megjelenésű), hanem dísznövényként is használják (Wu et al 2009).
Termőképesség oldalrügyeken és a termésminőség fokozása
A termesztők számára az egyik legfontosabb tulajdonság az oldalrügyeken való terméshozási képesség, mellyel a csak csúcsrügyből termő fajtákhoz képest fokozni tudjuk a termésmennyiséget. E nemesítési cél elérése érdekében a Pedro fajtát vonták be Magyarországon a keresztezéses nemesítési programban apai szülőként, mely további előnye, hogy világos színű és sima felületű terméshéjjal és világos színű magbéllel rendelkezik.
Néhány újabb nemesítésű fajtánál megjelent a fürtös terméshozási típus, mely azt jelenti, hogy a csúcsrügyekből nem egy termés, hanem 5 és 12 termésből álló, szőlőfürtre emlékeztető „terméscsoport” fejlődik ki. Általában a termések a fürtben kisméretűek, így csak igen kevés államilag minősített fürtös terméshozási típusú fajta található meg a diótermesztő országok fajtaválasztékában. Pozitív példaként említhetjük a szerb nemesítésű Tisza fajtát, mely fürtjei 7–9 termés tartalmaznak, melyek 30–33 mm átmérővel rendelkeznek. A fürtös terméshozási típusú diófajták további hátránya az alacsony fánkénti termésmennyiség. A példaként említett ’Tisza’ ez alól is kivétel, mivel bő termőképességgel rendelkezik.
A kaliforniai diónemesítési programban a termőképesség fokozása érdekében a Payne fajtát használták egyik szülőként, melynek eredményeként állították elő a Vina, Serr, Howard, és Chandler fajtákat (Ramos, 1998). Napjainkban három újabb fajtát (Sextont, Gillet, Forde) emeltek ki a nemesítés eredményeként, melyek terjedése néhány éve kezdődött meg. Mindegyik újonnan nemesített fajta kevésbé fogékony a dió xantomónászos betegségével szemben, oldalrügyeken is terem, bőtermő, korai érési idejű, jó bélminőséggel rendelkezik.
Az utóbbi időben egyre nagyobb igény mutatkozik piros belű diófajták iránt. Egyelőre csak néhány piros magbelű fajta található meg a fajtaválasztékban. Európában nincs ilyen fajta a termesztésben, csak génbanki gyűjteményekben őrzik a nemesítők. Kaliforniában is csak a Robert Livermore az egyetlen piros magbelű fajta, melyet egyelőre kis felületen termesztenek (McGranahan és Leslie 2004).
A diót károsító kórokozókkal kapcsolatos kutatások
Dió esetében a Xanthomonas arboricola pv. juglandis illetve a BAN (Brown apical necrosis magyar néven a dió csúcsi barnulása) kórokozókkal szemben toleráns vagy ellenálló genotípusok nemesítése a világ különböző nemesítési műhelyeiben elkezdődött. A BAN betegséget a Xanthomonas arboricola pv. juglandis és a Gnomonia leptosyla, valamint egyéb Alternaria és Fusarium fajok együttesen okozzák. Mindkét betegség elleni hatékony szabadföldi védekezés kidolgozása jelenleg is folyik. Ebbe a munkába kapcsolódott be a BCE Gyümölcstermő Növények Tanszéke és az Érdi Kutató, mely során a legnagyobb magyarországi dióültetvényben végeztek felméréseket mindkét betegség terjedését és az ellenük irányuló kémiai védekezés hatékonyságát illetően.
A kutatók egyetértenek abban, hogy a Xanthomonas előfordulása szorosan összefügg a diófák fiziológiai állapotával, ezen belül is a víz- és tápanyag-ellátottságával. Azokban az ültetvényekben, ahol kedvezőtlen a víz- és tápanyag-ellátottság, illetve gyakori száraz periódusok vannak, nagyobb mértékben terjedt a Xanthomonas az optimálisan ellátott ültetvényhez képest (Parveaud et al. 2009). Chevallier et al (2009) szerint a különböző diófajták Xanthonomas baktériummal szembeni viselkedése nagymértékben függ hidroxifahéjsav-származék és flavonoidtartalmuktól. Mindkét vegyületet fertőzés esetén csak akkor termeli a diófa, ha megfelelő vízellátottsággal rendelkezik a vegetációs idő során.
Kísérleti eredmények alapján a Xanthomonas arboricola pv. juglandis baktérium a virágzáskor 103 és a 107 közötti koncentrációt ér el, ezért célszerű ekkor elkezdeni a vegyszeres védekezést (Giraud et al 2009). Innovatív védekezési megoldás Chevallier és Laymajoux szerint (2009), ha a védekezéshez szükséges réztartalmú hatóanyagot kelát formájában juttatjuk ki, így kisebb rézmennyiséget használunk fel hatékonyabban. Hatékony megoldás Jenkins és munkatársai (2009) eredményei alapján, ha bakteriofág törzsekkel védekezünk e kórokozó ellen. Süle és munkatársai (2007) felhívják a figyelmet arra, hogy a különböző Xanthomonas arboricola pv. juglandis törzsek egymástól eltérőn viselkednek a különböző hatóanyagú és koncentrációjú baktericid készítményekkel szemben.
A Xanthomonas mellett egyes mediterrán klímájú országokban nagy problémát okoz az Agrobakterium tubefaciens, mely ellen nem toleráns az interspecifikus Paradox alanyfajta. Az újabb kutatási eredmények szerint a saját gyökéren előállított Chandler, Vina és Serr diófajták kevésbé fogékonyak e kórokozóval szemben Kaliforniában (Hasey et al 2009).
A dió fajtahasználata Magyarországon
A KSH 2001-es adatai szerint a magyar dióültetvények 40%-át a Milotai 10 fajta adta, melyet a 21%-os részesedéssel követett az ’Alsószentiváni 117’. A harmadik legnagyobb mértékben telepített diófajta a magyar termesztésben a Tiszacsécsi 83, mely termesztési arány 7%.
Az újonnan nemesített hibrid diófajták iránt jelentős igény mutatkozik a termesztők részéről, de terjedésük nehézkes. A hibridfajták termesztése a termesztéstechnológia pontos betartását igényli. Tény, hogy csapadékos időben a Xanthomonas arboricola pv. jugladis nagymértékben fertőzi a hibrid diófajtákat. A termesztők elismerik viszont az oldalrügyből is termő diófajták hagyományos fajtákhoz képest nagyobb termésmennyiségét és a közel megegyező termésminőségüket.
A külföldi diófajták aránya 27% a magyar termesztésben. Ennek az óriási aránynak az oka abban keresendő, hogy a szükséges oltványmennyiséget nem minden esetben tudták biztosítani a hazai faiskolák.
Fajtaújdonságok – az új ültetvényekbe ajánlott új hazai és külföldi fajták
A külföldön nemesített diófajták közül olyan ígéretes fajtákat mutatunk be ebben az alfejezetben, melyek genetikai hátterük alapján, illetve megfelelő adaptációs képességük esetén Magyarországon is sikeresen termeszthetők.
Ukrán fajták
Csernivetszky 1
Nőelőző virágzású, virágzása későn, májusban kezdődik. Termései szeptember második dekádjában érnek. Termése ovális alakú, csúcsa hegyes, átlagos méretű, tömege 11–13 g. A héja vékony (0,9 mm), majdnem sima felületű, könnyen törhető. A dióbél aránya 50–55%, magbelének színe világos, íze jó. Korán termőre fordul, termése és levele ellenáll a diót károsító kórokozóknak. Fája közepes növekedésű (Zatokovy 2009).
Ukrán kutatókkal közösen végzett érzékszervi bírálat során arra az eredményre jutottunk, hogy a Csernivetszky 1 fajta magbele kiváló tulajdonságokkal rendelkezik. A felkért bírálók magasabb pontszámot adtak a ’Csernivetszky 1’ magbelére, mint a Milotai 10 kontrollfajtáéra.
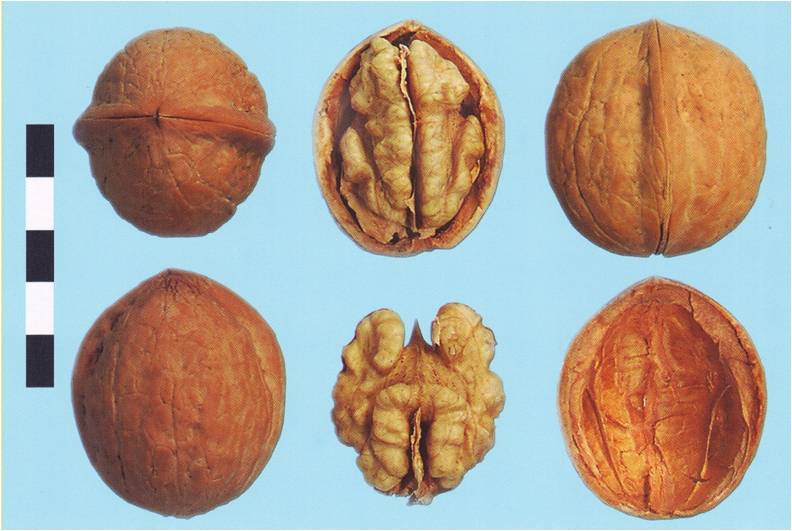
Cserniveszky 1 diófajta (fotó: Zatokovy)
Bukovynszky 1
Hímelőző virágzású fajta. Termése átlagos méretű, tömege 10–14 g, kerekded, hegyes végű. Termésének héja vékony (0,8 mm), majdnem sima felületű, könnyen törik. A dióbél aránya átlagosan 52,4%, magbele világos színű, könnyen törhető, íze jó. Kiemelkedő magas terméshozama és termésminősége. Fája közepes növekedési erélyű (Zatokovy 2009). A Bukovynszky 1 fajta elsősorban szárított héjas formában közelíti meg a ’Milotai 10’ áruértéket.

Bukovynszky 1 (fotó: Zatokovy)
Klishkivszky
Virágzása nőelőző, virágzásának kezdete későn, május elején van. Érési ideje szeptember közepe. A zöld burok – a többi diófajtától eltérően - a termés csúcsánál kezd repedni. Termése hosszúkás alakú, átlagos méretű, tömege 11–13 g. A héj vastagsága 1,2 mm, sima felületű, sárgásbarna színű, szárított héjas állapotban áruértéket megközelíti a Milotai 10 fajtáét. Terméshéja könnyen törhető. A dióbél aránya 49–50%, színe világos, íze jó. Fája erős növekedésű (Zatokovy 2009).
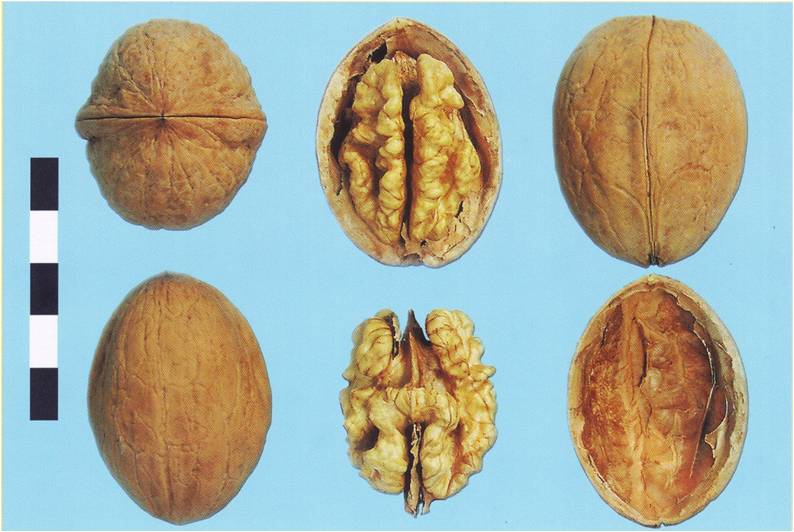
Klishkivszky diófajta (fotó: Zatokovy)
Grozynetszky
Hímelőző virágzású, a virágzás kezdete május eleje. Termései szeptember végén érnek. Termései oldalirányban hasadnak, nagyméretűek, trapéz alakúak, tömegük 14–15 g. Terméshéja vékony, 0,9 mm, kissé barázdált felületű, könnyen törik. Terméshéjának minősége az Alsószentiváni 117 fajtához hasonló. A dióbél aránya 49–53%-, színe világos, íze jó. Fája közepes növekedésű. (Zatokovy 2009).

Grozynetszky diófajta (fotó: Bujdosó)
Szlovén diófajták
Zdole-62
Késői virágzású fajta. Közepes, de rendszeres terméshozamú. Termése lapított trapéz alakú, nagyméretű. Termésének tömege 9,5–11 g, bélaránya 41–47%. Közepesen barázdált vagy sima felületű a héja, világos színű. Bélszíne világos, könnyen tisztítható. Fája középerős növekedésű, felfelé törő. Erősen fogékony a betegségekre. (Solar és Stampar 2005).

Zdole-62 fajtajelölt (fotó: Bujdosó)
Erjavec
Nőelőző virágzású fajta, nővirágai középidőben nyílnak. Termőképessége jó, terméseinek egy részét oldalrügyein fejleszti, rendszeresen és bőven terem. Termése gömbölyded alakú, kicsi - közepes méretű, tömege 8,6–10,2 g, bélaránya 49–54%. Héj felülete sima, a világos és a középbarna közötti átmeneti színű, bélszíne világos, könnyen tisztítható. Fája erőteljes növekedésű, koronája szétterülő. Nagyon fogékony a bakteriális betegségekre. (Solar és Stampar 2005).

Erjavec diófajta (fotó: Bujdosó)
Francia fajták
Franquette
Régi francia nemesítésű fajta, mely a francia diótermesztés sikerességét megalapozta. Fakadási ideje kései, de termesztésének sikerességét veszélyezteti, hogy hajtásai későn érnek be. Hímelőző virágzású, a barkák nyílása 10–12 nappal előzik meg a nővirágok nyílását. Érési ideje: október eleje-közepe. Termése 10–12 g tömegű, hosszúkás alakú, kissé rücskös felületű, világosbarna színű. (9.9. ábra) A héj varratai jól záródnak. Magbele kissé megnyúlt alakú, világosbarna színű, magbelének aránya 46%. Fája középerős-erős növekedési erélyű (Verhaeghe 2011). Jelenleg nincsenek tapasztalatok a Franquette fajta magyarországi termesztéséről.

Franquette fajta (fotó: Internet1)
Fernor
Új nemesítésű francia fajta, melyet a ’Franquette’ és a ’Lara’ keresztezésével állítottak elő. Kései fakadási idejű, korán termőre fordul, terméseinek nagy részét nagy arányban oldalrügyeken fejleszti. Érési ideje kései, szeptember vége – október eleje. Termése nagy, világos színű, magbelének szép színét hosszú ideig megőrzi, könnyen tisztítható, és jól tárolhatók a termései. Kiváló ízű fajta. Termőkorban 4–5 t/ha szárított héjas termésmennyiségre képes francia ökológiai körülmények között, s itt ellenáll a Xanthomonas-nak (Verhaeghe 2011). Magyarországi ökológiai körülmények között eddig jól vizsgázott. Magbelnek szalmasárga színét 9–12 hónapig megtartja.

Fernor diófajta (fotó: Internet 1)
Szerb fajták
Sampion
Hímelőző virágzású, az egyik legjobb hidegtűrésű fajta, kontinentális körülmények között is ajánlott a termesztése. Termése az ’Eszterházy I’-re hasonlít, igen szép, átlagtömege 14 g. A bél szép világos. Bélaránya 58%. Közepes növekedésű, igen bőtermő. Fogékony a Gnomonia-ra. Középerős növekedési erélyű, jó télálló és hidegtűrő, szerb adatok szerint kiválóan tűri a téli nagy hőingásokat. (Internet 2, Cerovic et al 2010).

Sampion diófajta (fotó: Internet 2)
Szrem
Hímelőző virágzású, bőtermő fajta. Termése nagyméretű, 14–15 g tömegű. Héja sima felületű és vékony, magbele világos színű, tetszetős. Bélaránya 57%. A diót károsító főbb betegségekkel szemben ellenálló. Közepes növekedésű, átlagos termőképességű. Középerős – erős növekedési erélyű. (Cerovic et al 2010., Internet 3)

Szrem diófajta (fotó: Internet 3)
Tisza
Fürtös típusú, bőtermő fajta. A fürt akár 20 nővirágot is tartalmazhat, azonban ebből 7–8 szokott teljes mértékben kifejlődni. Legtöbb évben homogám virágzású. Termése 15 g tömegű, kerek alakú, melyet a bél nem tölti ki egészen. Bélaránya a vékony héj miatt 51%. Különösen olyan termőhelyekre ajánlható, ahol eredményesen lehet szőlőt termeszteni. (Cerovic et al 2010., Internet 2).
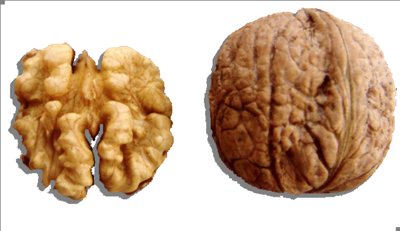
Tisza diófajta (fotó: Internet 4)
Bácska
A Sejnovo-val egyidőben fakad, és vegetációját 10 nappal korábban fejezi be. Hímelőző virágzású fajta. Közepes növekedési erélyű, igen bőtermő. Egyes oldalrügyeiből is terem. Termése 12 g tömegű, kúpos alakú, világos színű, vékony héjjal. A bél világos színű. Magbél aránya 52% körüli, olajtartalma 68–70%. Középerős növekedési erélyű (Cerovic et al 2010).

Bácska diófajta (fotó: Internet 2)
Bolgár fajták
Sheinovo
Szelektált magonc, Fakadási ideje középkorai. Virágzási ideje alatt tökéletesen mutatja a dichogámiát, virágzási jellege nőelőző. Érési ideje szeptember 2. illetve 3. dekádja. Terméseinek 30–40%-át oldalrügyeiből fejleszti. Termése középnagy, tömege 12,5 g. Magbelének színe borostyánsárga, bélaránya 54%. Fája erős növekedési erélyű. Jó ellenállóképességet mutat a xantomónászos betegséggel szemben, a gnomóniás betegséggel szemben viszont mérsékelten fogékony (Dzhuvinov et al 2010).

Sheinovo diófajta (fotó: Gandev)
Silistrenski
Szelektált magonc. Hímelőző virágzású. Termései szeptember közepén érnek. Terméseinek 30–40%-át oldalrügyein fejleszti. Termése középnagy, 11,5 g tömegű, 51,5% magbél tartalommal. Fája középerős növekedési erélyű. Jó ellenállóképességet mutat a xantomónászos betegséggel szemben, a gnomóniás betegségre viszont mérsékelten fogékony (Dzhuvinov et al 2010).

Silistrenski diófajta (fotó:Gandev)
Amerikai fajták
Chandler
A legnagyobb arányban termesztett hibrid diófajta a világon. Fakadási ideje kaliforniai körülmények között kései, hazánban középkorai. Terméseinek nagy részét oldalrügyeiből fejleszti, bár fiatal korban ez még nem nagyon jellemző. Érési ideje szeptember utolsó dekádja. Termései 13 g tömegű, 28–30 mm átmérőjű, sima héjfelületű, világos héj- és bélszínnel rendelkezik. Magbélaránya 49%. Fája középerős növekedési erélyű, félig feltörő habitusú. A xanthomónászos betegségre nagyon fogékony (Hendricks et al 1998). Eddigi tapasztalataink alapján Magyarországon védett termőhelyen eredményesen termeszthető, bőtermő fajta, de termésmérete öntözetlen körülmények között 25 mm körüli.

Chandler diófajta (forrás: Internet 3)
Román nemesítésű diófajták
Valrex
Virágzási ideje korai (április utolsó dekádja), nőelőző virágzású fajta. Érési ideje korai (szeptember első dekádja). Termései ovális alakúak, átlagosan 15,3 g tömegűek, terméshéjuk finoman erezett. Magbelének aránya 51%, könnyen törhető és tisztítható fajta. Magbele kiváló ízű és világos színű. Fája középerős növekedési erélyű, korán termőre forduló, bőtermő. Közepesen fogékony a diót károsító gombás és baktériumos betegségekre. Nagyon jó téltűrő képességgel rendelkezik. Terméseit csak csúcsrügyből fejleszti (Botu et al 2010/a, 2010/b, Cosmulescu 2010/a, 2010/b, 2012).

Valrex diófajta (fotó: Botu)
Jupâneşti
Virágzási ideje középkései (május 6. és 10. közé esik), nőelőző. Érési ideje szeptember második dekádjában van. Termése elliptikus alakú, átlagosan 12,2 g tömegű, finoman erezett terméshéjú. Béltartalma 49%, termése könnyen törhető és magbele könnyen tisztítható. Magbele kiváló ízű, világos borostyán színű. Fája erős növekedési erélyű, későn termőre forduló, de később bőtermővé válik. A diót károsító valamennyi kórokozóval szemben jó ellenállóképességet mutat, jó téltűrő képességű, csak csúcsrügyből termő fajta (Botu et al 2010/a, 2010/b, Cosmulescu 2010/a, 2010/b, 2012).

Jupâneşti diófajta. (fotó: Botu)
Valcor
Virágzási ideje korai (április 20. és 30. közötti Románában), egyes évjáratokban nővirágai a középkorai időszakban (május 1. és 5. között) nyílnak. Virágzásának jellege hímelőző. Érési ideje középkorai, termései szeptember második dekádjában érnek. Termése széles ovális alakú, átlagosan 14,3 g tömegű, finoman erezett. Bélaránya 52%, könnyen törhető és tisztítható fajta, magbele világos színű. Fája erős növekedési erélyű, korán termőre forduló, bőtermő, közepesen fogékony a diót károsító baktériumos és gombás betegségekre, nagyon jó téltűrő képességgel rendelkezik. Terméseit csak csúcsrügyből fejleszti (Botu et al 2010/a, 2010/b, Cosmulescu 2010/a, 2010/b, 2012).

Valcor diófajta. (fotó: Botu)
Valmit
Virágzási ideje korai (április harmadik dekádja Romániában), nőelőző virágzású fajta. Érési ideje szeptember második dekádja. Termése gömbölyű, átlagosan 12,1 g tömegű, terméshéja finoman erezett felületű. Magbelének aránya 53%, könnyen törhető és tisztítható fajta. Magbele kiváló ízű és világos színű. Fája erős növekedési erélyű, bőtermő fajta, Közepesen fogékony a baktériumos betegséggel szemben, kevésbé fogékony a diót károsító gombás betegségekre. Terméseit csúcsrügyből fejleszti (Botu et al 2010/a, 2010/b, Cosmulescu 2010/a, 2010/b, 2012).

Valmit diófajta (fotó: Botu)
Magyarországi termesztésre nem javasolható diófajták
A külföldön nemesített diófajták kiválasztása során elsődleges szempont a fajták megfelelő alkalmazkodóképessége a magyar ökológiai adottságokhoz. E szempont alapján teljes mértékben kizárhatók a mediterrán klímaviszonyok között nemesített török, iráni és a kaliforniai fajták döntő többsége (a ’Pedro’ és a ’Chandler’ kivételével) a magyarországi termesztésből, mivel nem rendelkeznek hazánk klímaviszonyainak megfelelő téltűrő- és télálló-képeséggel.
Az európai országokban nemesített diófajták közül is számos fajta magyarországi termesztése nem lehetséges, ennek oka elsősorban a korai fakadási idejükkel magyarázható. Ez azt jelenti, hogy célszerű az április 20-a előtt fakadó fajták magyarországi termesztéséről lemondani. Továbbá kerülni kell a hegyvidéki, hűvös, párás, kiegyenlített klímaviszonyokból származó fajták (egykori csehszlovák fajták) termesztését is Magyarországon, mivel ezek a fajták hazánk arid klímája miatt sötét, eladhatatlan bélminőséget produkálnak. Mellőzni kell a dió xantomónászos betegsége iránt nagymértékű fogékonyságot mutató fajták honosítását is, mert termesztésük költséges, bélminőségük minden esetben jelentősen függ az évjárattól. Toleráns/rezisztens fajták esetében az adaptációs kísérletek megkezdése előtt tudni kell a kórokozó pontos adatait (rassz), amivel szemben ellenálló-képességet mutat a termeszteni kívánt fajta. Limitáló tényező még az érési idő is. A fajtahasználati trendeknek megfelelően nem kifizetődő október közepe utáni érési idővel rendelkező fajta termesztésével foglalkozni, mivel ebben az időszakban a termések beérése kockázatos illetve a piac telített.
Gesztenye ( Castanea sativa Mill.)
A gesztenye nemesítésének rövid története
A Castanea genus Eurázsiában és a Kaukázusban honos, déli elterjedtsége a 37. északi szélességi köréig terjed, egészen Tunézia, Szíria illetve Libanon partvidékéig a Föld északi féltekéjén. Az ókori görögök és rómaiak hódításai révén az európai gesztenye (Castanea sativa Mill.) areája a Földközi-tenger partvidékéről az Ibériai-félszigetre, valamint Európa középső és északabbra fekvő részeire (Diamandis 2009) szélesedett ki.
Ott, ahol a természetes gesztenyeállománynak nagy a genetikai variabilitása tájszelekcióval állították elő az első nemesített gesztenyefajtákat. Oroszország nyugat-kaukázusi régiójában igen nagyszámú Castanea populáció található, mely összefüggő erdőket alkot. E nagyszámú populáció változatos genetikai hátteret ad az erdészeti és a kertészeti célú nemesítő munkához (Pridnya et al. 2009).
Magyarországon az Állami Gyümölcs- és Dísznövénytermesztési Kutató-Fejlesztő Közhasznú Nonprofit Kft. jogelődjeinél Szentiványi Péter irányításával folyt gesztenyenemesítés, melynek eredményeként a magyarországi gesztenyepopulációból szelektálta a Kőszegszerdahelyi 29, Iharosberényi 2, Iharosberényi 29, Nagymarosi 22, Nagymarosi 37 és Nagymarosi 38 fajtákat. A tudatos fajtahasználat azonban csak az 1960-as évek végétől kezdődött meg hazánkban, mivel addigra dolgozták ki a gesztenye szaporításának faiskolai technológiáját (Szentiványi 1988). Szlovéniában is jelentős gesztenye populáció található, melyből ígéretes fajtákat, illetve genotípusokat szelektáltak a Ljubljanai Egyetemen (Solar és Stampar 2009). Kínában is nagy genetikai változatosság figyelhető meg, ezért nagymértékben használják a tájszelekciót, mint nemesítési módszert a gesztenyenemesítők (Cheng és Huang 2009). Továbbá tájszelekciót folytatnak Bulgáriában (Bratanova-Doncheva et al. 2009) és Albániában is (Lushaj és Tabaku 2009).
Számos kutatóhelyen az utóbbi két-három évtizedben indult meg a keresztezéses nemesítés, mely elsősorban fajhibridek előállítását eredményezte. A keresztezéses nemesítés során a Castanea sativa Mill. x Castanea mollissima Blume (kínai gesztenye) vagy a Castanea sativa Mill. x Castanea crenata Siebold & Zucc. (japán gesztenye) kombinációkat használták (Botu 2009). E kombinációkból származó interspecifikus hibrideket használnak ma is Braziliában (Yamanishi et al. 2009). Az Amerikai Egyesült Államokban az európai gesztenye és a kínai gesztenye közötti fajhibridek nemesítésével foglalkoznak, mivel a természetes gesztenyeállományt alkotó Castanea dentata-t Borkh. a gesztenyerák szinte teljesen tönkretette.
A biotechnológiai módszerek közül említésre méltó, hogy mikroszatellit markerezéssel Hozova és munkatársai (2009) a legidősebb csehországi természetes gesztenyepopuláció diverzitását vizsgálják.
Fő nemesítési célok
Gesztenye esetében kiemelt nemesítési cél a bőtermőképesség, a korai érési idő, az egyöntetű érés és a kiváló termésminőség, mely nagy termésméretet, kerekded, telt termésalakot, sötét termésszínt, egyedi, jellegzetes ízt, alacsony zsírtartalmat jelent. Fontos értékmérő tulajdonság a magbél tisztíthatósága, masszakészítésre való alkalmasság és egyöntetű állománya (ne legyen „lyuk” a magbél közepén) is. A terméstulajdonságok mellett lényeges a zöld burok egyöntetű felnyílásának ideje. Az öntermékenység kérdése rendszeresen felmerül a gesztenyenemesítő műhelyekben (Soylu és Mert 2009, Serdar et al. 2009, Pandit et al. 2009, Solar és Stampar 2009). Napjainkban előnybe részesítik a maróni típusú gesztenyét, melynek kupacsában egy nagyméretű termés található.
Ezen kívül nagyon fontos a gesztenyét károsító kórokozókkal szembeni ellenálló-képesség, ezen belül is kiemelt nemesítési cél a Cryphonectria parasitica Murr. Barr. (syn.: Endotia parasitica), Phytophthora cambivora és Cytospora spp. kórokozók elleni nemesítés. A rezisztencianemesítésbe számos ország helyi populációból kiemelt genotípusokat von be (Soylu 2009, Celiker és Onogur 2009).
Anagnostakis és munkatársai (2009) felhívják a figyelmet a gesztenye gubacsdarázs (Dryocosmus kuriphilus) elleni rezisztencianemesítés fontosságára. Az áltata vizsgált populációban a gesztenye gubacsdarázzsal nem fertőzött genotípusok citoplazmájában feltehetőleg egy monogenikus domináns gén található. Sartor és munkatársai (2009) Olaszországban végzett vizsgálatai során Castanea sativa × Castanea crenata fajok keresztezésével nemesített hét hibridfajta Dryocosmus kuriphilus-szal szembeni fogékonyságát vizsgálta, melyek közül egyedül a Bouche de Bétizac fajtán nem okozott kárt a gesztenye gubacsdarázs.
Fontos a gesztenyenemesítésben az újonnan nemesített genotípusok esetében a gyenge vagy középerős növekedési erély, felfelétörő koronahabitus erős, közel függőlegesen álló koronaelemekkel az egyszerűbb gépi betakaríthatóság miatt (Soylu és Mert 2009).
Termesztéstechnológiai szempontból fontos a gesztenye genotípusok szaporíthatóságának illetve betakarításra való alkalmasságának vizsgálata (Pandit et al. 2009).
Hipovirulens törzsekkel (gyengített virulenciával rendelkező Criphonectria parasitica törzsekkel) tudunk a leghatékonyabban „védekezni” a Criphonectria parasitica ellen. Azonban a hipovirulens törzsek állandó vizsgálatára és a laboratóriumi tesztelésére van szükség, mivel nem minden hipovirulens törzs kompatibilis a fertőző törzsekkel (Vidóczi et al 2007). Jelenleg Magyarországon a Nyugat-Magyarországi Egyetem Erdőmérnöki Karán található a magyar gesztenye hipovirulens törzsek gyűjteménye. A Debreceni Egyetemen Magyarország, Szlovákia, Ukrajna és Románia gesztenye kéregrák törzseinek felmérése folyik (Radócz et al. 2009). Jelenleg a világ egyetlen gesztenyetermesztő országában sincs tökéletes növényvédelmi technológia a gesztenyerák ellen (Döken 2009). Svájcban sikeresen használják a CHV-1 hipovirulens törzset, mellyel kezelt növényi részek jól viselkednek a kezelést követő újrafertőzések után is (Heininger és Ringling 2009). Olaszországban a nagyobb gesztenyepopulációkban „természetes úton” terjedő hipovirulens törzsek jelenthetik a megoldást a Criphonectria parasitica elleni küzdelemben. Horvátországban négy populáció (Markuevac 1, Markuevac 2, Samobor, Sljeme) vizsgálatát végezték el, melyekben hat hipovirust tartalmazó mintát találtak (Krstin et al. 2009).
A Phytophthora cambivora ellen a Trichoderma viride és a Trichoderma harzianum jelentik a biológiai védekezés alapját (Turchetti és Maresi 2009).
A nemesítés genetikai forrásai
Az európai gesztenyenemesítés a Castanea sativa (2n=24) fajon alapszik, az európai gesztenyetermesztés géncentruma Törökországban található. A török gesztenyetermesztés központjai Anatólában, Marmara és Égei régiókban valamint a Fekete-tenger környékén vannak. Törökországban nagymértékben használják a tájszelekciót, illetve gyűjtött genotípusokat használnak fel a rezisztencianemesítésben (Soylu 2009, Soylu és Mert 2009, Serdar et al. 2009). Emellett nagy jelentősége van az interspecifikus fajhibridek előállításának, mivel a Castanea sativa jó génforrás, de fogékonyabbak gesztenyét károsító Criphonectria parasitica kórokozóra, a Castanea mollissima Bl. (kínai gesztenye) és a Castanea crenata Sieb. és Zucc. (japán gesztenye) fajok viszont ellenállóképeséggel rendelkeznek e kórokozóval szemben
Kínában a Castanea henryi is felhasználják a nemesítési programban mérsékelt növekedési erélyű, Marroni típusú fajták előállítására.
Az Amerikai Egyesült Államokban a Castanea dentata őshonos. Sajnálatos módon a Castanea dentata természetes állományát a Criphonectria parasitica szinte teljes egészében kipuszította, ezért jelenleg főleg az európai gesztenye és a japán gesztenye fajhibrideire épül az USA gesztenye szortimentje (Fullbright et al. 2009).
Szlovákiában a Castanea sativa × Castanea crenata fajokkal végzett keresztezéses nemesítés során keletkezett hibridek kisebb különbségeket mutattak a gesztenye kéregrákkal szemben (Bolvansky et al. 2009).
Japánban a japán gesztenyén (Castanea crenat) alapszik a nemesítés. E gesztenyefaj jó nemesítési alapnak bizonyul, mivel nagyobb méretű terméseket hoz, mint a Castanea mollissima (kínai gesztenye). Igaz azonban az is, hogy a japán gesztenye nehezebben hámozható, illetve tisztíható a kínai gesztenyéhez képest (Takada et al. 2009).
A gesztenye fajtahasználata Magyarországon
A KSH 2001-es adatai szerint Magyarországon összesen 467 ha gesztenyeültetvény található. Legnagyobb mennyiségben egyéb, a Nemzeti Fajtalistán nem található fajtákat termesztünk. Feltehetően az „egyéb” megjelölés magoncokat, illetve kisebb arányban külföldi nemesítésű fajtákat takar. Magyar nemesítésű, Nemzeti Fajtalistán lévő fajták közül legnagyobb mértékben a Kőszegszerdahelyi 29 (termesztési részaránya 36,2%) fajtát termesztjük. A Kőszegszerdahelyi 29 fajtát kisebb arányban követ a többi fajta: az Iharosberényi 29 (termesztési részaránya 7,1%), Iharosberényi 2 (termesztési részaránya 6,7%), Nagymarosi 22 (termesztési részaránya 4%), Nagymarosi 37 illetve Nagymarosi 38 (termesztési részarányuk 2,2 – 2,2%) fajták.
Fajtaújdonságok – az új ültetvényekbe ajánlott új külföldi fajták
Marrone di Marradi
Termései igen nagyméretűek, középbarna színűek, csíkozottak, fénylő felületűek. Népszerűségét nemcsak nagy termésméretének és nagy béltartalmának, hanem kiváló élelmiszeripari felhasználhatóságának is köszönheti (Internet1).

Marrone di Marradi (forrás: Internet1)
Marrone di Castel del Rio
A földrajzi eredetvédelemmel rendelkező fajtát a Santero völgyből szelektálták. Termésmérete igen nagy, héja sötétbarna színű, csíkozott. Magbele kiváló édes ízű, sokoldalúan felhasználható (Internet2).

Marrona di Castel del Rio (forrás: Internet2)
Kozjak-1
A Dráva völgyében a kozjaki lejtőkön (Szlovénia) szelektált fajta. A szlovén fajták közül a legnagyobb termésmérettel rendelkezik. Terméseinek átlagtömege 16,4 g (61 termés/kg). Termése széles ovális alakú, sötétbarna színű, hosszirányban csíkozott. Pecsétje hosszú és széles. Termése mindig egy embriót tartalmaz (Solar és Stampar 2009).
Avbar (syn.: MP-30)
Szlovén nemesítésű tájszelekcióval nemesített fajta. Termése középnagy, 13,7 g tömegű (73 termés/kg). Termése kerekded vagy elliptikus alakú, széles és nagy pecséttel rendelkezik. Termése vöröses vagy sötétbarnás színű, esztétikusan csíkozott (Solar és Stampar 2009).
L-voda
Szlovén nemesítésű tájszelekcióval nemesített fajta. Termése nagyméretű, átlagosan 15,3 g tömegű (65,3 db/kg). Termése széles elliptikus alakú, széles és nagy pecséttel rendelkezik. Terméshéja vörösesbarna, nem csíkozott (Solar és Stampar 2009).
Mogyoró ( Corylus avellana L.)
A mogyoró nemesítésének rövid története
A hazánkban termesztett közönséges mogyoró (Corylus avellana L.) Közép-Európában, a Balkán-félszigeten, Nyugat- és Kelet-Ázsiában és Észak-Amerikában is őshonos (Tomcsányi 1979). Ennek ellenére Magyarországon csak igen kis felületen, védett mikrokörzetekben termesztjük. Hazánkban csak olyan fajták termeszthetők sikeresen, melyek a hosszú mélynyugalmi időszakot elviselik, kései virágzási idővel rendelkeznek.
Az első nemesített mogyorófajták magoncok voltak, melyeket később felváltották a tájszelekcióval, majd a keresztezéses nemesítéssel előállított fajták. A keresztezéses nemesítés igen népszerű az Amerikai Egyesült Államokban, Olaszországban, Franciaországban és Romániában. Emellett minden mogyorótermesztő országban nagymértékű honosítás figyelhető meg. Napjainkra az olasz és amerikai nemesítésű fajták előretörése jellemző.
Az Állami Gyümölcs- és Dísznövénytermesztési Kutató-Fejlesztő Közhasznú Nonprofit Kft. jogelődjeiben folyt mogyorónemesítési program, Szentiványi Péter tájszelekcióval előállította a Bőtermő nagy fajtát. A tájszelekcióból adódóan a ’Bőtermő nagy’ igen jól alkalmazkodik a hazai ökológiai körülményekhez, melynek eredményeként napjainkra jelentős mértekben elterjedt ez a fajta az országban (KSH 2003). A ’Bőtermő nagy’ mellett szintén helye van a magyar termesztésben, a Kertészeti és Élelmiszeripari Egyetemen szelektált Nagy tarka zelli K. 5 fajtának, mely egy ismeretlen származású külföldi fajta hazánkban szelektált genotípusa.
A magyar nemesítésű fajták mellett szinte valamennyi jelentősebb világfajta is megtalálható a mogyoró hazai fajtaszortimentjében. Igaz azonban, hogy valamennyi világfajtából magyar ökológiai körülmények között szelektált klónfajtákat termesztünk (Cosford K.2, Római mogyoró K. 1, Grosse aus Trient K. 3, Bollwilleri csoda K.4, Princess Royal K. 6). Nagyon sok esetben nem egyszerű megállapítani a fajták eredetét, mivel igen gyakran jönnek létre termesztett fajták között természetes hibridek.
A mogyoró termesztése egyre inkább terjed a világon, ezt bizonyítja az a tény is, hogy 1997-ben míg csak 24, 2009-ben már 30 országban volt jelentős mogyorótermesztés a FAO adatai szerint. A mogyorót termelő országok számának növekedése ellenére ezt a gyümölcsfajt továbbra is mediterrán klímaviszonyok között termesztik, a világ össztermésének 80%-át Törökország és Olaszország adja (Fidigelli és Salvador 2009). A mediterrán klímájú országoktól északra fekvő országokban a mogyoró a kockázatosan termeszthető gyümölcsfajok közé sorolható, a mogyoróültetvények terjedését azonban generálhatja a nyári szarvasgomba (Tuber aestivum Vittadini) iránti élénk érdeklődés, mivel a mogyoró igen jó gazdanövénye e fajnak.
Fő nemesítési célok
Mogyoró esetében az egyik legfontosabb nemesítési cél a világ minden táján a termésbiztonság, ehhez kapcsolódóan a kései virágzási idő. A mogyoró virágai nyílnak a legkorábban a termesztett gyümölcsfajok közül, ezért különösen fontos a kései virágzási idő Közép-Európában is.
Fontos a bőtermőképesség is, egyes ukrán kutatók szerint termőkorban 8 kg/bokor termésmennyiséget kell elérnie az ígéretes fajtáknak.
Érési idő tekintetében lényeges a korai érési idő az elsődleges cél, így a termesztők hamar meg tudnak jelenni a piacon friss áruval.
Nagyon fontos a kiváló termésminőség. A különböző mogyorótermés alakok közül a kerek vagy gömbölyded termésalakok preferáltak, mivel kiváló termésméretet érnek el. Ezen kívül fontos még a kedvező, világos szín, vékony terméshéj, jó íz és a legalább 43–50%-os béltartalom. Számos minősített mogyorófajta esetében problémát okoz heterogén termésminőség.
Egy jó mogyorófajta esetében lényeges a jó téltűrőképesség, mely nagyon sok termőhelyen a termesztés sikerének kulcsát jelenti. Emellett a szárazságtűrés is előtérbe kerül a nemesítési célok között.
Sarjhozam tekintetében a gyenge sarjhozam a legkedvezőbb, mivel így hatékonyabban tudunk védekezni a Phytoptus avellanae ellen (Slyusarchuk és Ryabokon 2005, Rovira et al 2005, Fidigelli és Salvador 2009).
A nemesítés genetikai forrásai
Az európai mogyorótermesztésben kizárólag a Corylus avellana (2n=22) fajt termesztjük (Thompson et al. 1996). Ukrajnában a téltűrőképesség fokozása érdekében a Corylus heterophylla Fisch. ex Trautv fajt vonták be a nemesítő munkába (Slyusarchuk és Ryabokon 2005). Grúziában a Corylus pontica és a Corylus conchica fajokra épül a nemesítés és a termesztés is (Mirotadze 2005).
A kínai mogyorónemesítésben a Corylus heteropylla és a Corylus avellana kombinációkat használnak, mivel a Corylus heterophylla jó tél- és fagytűrőképességgel, de szerény termésmennyiséggel és kisméretű (1 g alatti) termésekkel rendelkezik. A Corylus avellana termőképessége és terméseinek minősége azonban kiváló. E két faj keresztezéséből számos interspecifikus hibrid jött létre, melyek közül néhány már a termesztésben is megtalálható (Xie et al 2005).
Amerikai kutatók az Oregoni Állami Egyetemen az Anisogramma anomala (Peck) E. Müller kórokozó gombával szemben végeznek rezisztencianemesítést, mely eredményeként állították elő a Santiam fajtát (McCluskey et al 2009). Az ország keleti partvidékén a New Yersey-i Egyetemen is folyik rezisztencianemesítés, melyben a Zimmerman és a spanyol nemesítésű Ratoli fajtákat használják fel a rezisztencia érdekében, mivel mindkét fajta jól örökíti ellenállóképességét. (Molnar et al 2009).
A patogének ellen biotechnológiai módszerekkel állítanak elő rezisztens genotípusokat Pilloti és munkatársai (2009). Kutatásuk a rezisztens gének (R gének) megkeresésén, illetve a patogénekhez kapcsolódó molekuláris háttér tanulmányozásán alapszik.
A mogyoró fajtahasználata Magyarországon
Magyarországon az árutermelő mogyoróültetvények negyedét a Római mogyoró K.1. fajta teszi ki, melyet szorosan követ a magyar nemesítésű ’Bőtermő nagy’ (termesztési részaránya 21,8%), az angol nemesítésű ’Cosford’ (termesztési részaránya 18,5%) és a ’Bollwilleri csoda K.4’ (termesztési részaránya 14,45%). Figyelemre méltó, hogy az egyéb mogyorófajták aránya igen magas, közel 20% a magyar mogyorótermesztésben (KSH, 2003).
Fajtaújdonságok – az új ültetvényekbe ajánlott új hazai és külföldi fajták
A külföldön nemesített mogyorófajták közül olyan ígéretes fajtákat mutatunk be ebben az alfejezetben, melyek genetikai hátterük alapján illetve megfelelő adaptációs képességük után Magyarországon is sikeresen termeszthetők.
Barcelona
Régi amerikai nemesítésű, nagy alkalmazkodó-képeséggel rendelkező mogyorófajta, Nyugat-Európában igen elterjedt. Középidőben virágzik, középkésőn érő fajta. Kupacsa egyharmaddal hosszabb, mint a termés. Termése nagyméretű, közép- vagy sötétbarna színű, kerek alakú, középesen vastag terméshéjjal és 39–42%-os béltartalommal rendelkezik, jó ízű (1. ábra). Bőtermő fajta. Növekedési erélye erős, nagy bokrot nevel (Solar 2000, Internet 1).

Barcelona mogyorófajta (forrás: Internet1)
Istarski duguljasti
Horvát nemesítésű fajta, melyet egész Horvátországban termesztenek. Bőtermő fajta. Termései csövesek, nagyméretűek, jó ízűek. Középerős növekedési erélyű, felfelétörő habitusú bokrot nevel. Erősen sarjadzik (Miljkovic 2000).

Istarski duguljasti (Internet2)
Istarski okruglasti
Isztrián nemesített fajta, melyet mogyorótermesztő vidékeken termesztenek. Középidőben virágzik és érik, termőképessége közepes - nagy. Termései kerek alakúak, nagyméretűek, jó ízűek, 45%-os béltartalommal rendelkeznek. Erős növekedési erélyű, félig felálló szerkezetű bokrot nevel. Gyengén sarjadzik. (Miljkovic 2000)

Istarski okruglasti (Internet3)
Tonda Gentile Romana
Olasz nemesítésű fajta. Nővirágai középidőben vagy későn nyílnak, kevésszámú hímvirágzata középidőben nyílik. Érési ideje középidejű. Kupacs a termésen túlnyúlik. Termése gömbölyű alakú, középnagy, világosbarna színű. Magbele gömbölyű alakú, középnagy, egyöntetű, jó ízű. Magbelének aránya 57%. Bokra félig felálló szerkezetű, gyenge növekedési erélyű, közepesen sarjadzó (Internet4).

Tonda Gentile Romana (Internet4)
Ellenőrző kérdések:
Dió
1. Ismertesse a magyar diónemesítés történetét!
2. Melyek a diónemesítés legfontosabb céljai?
3. Melyek a dió legfontosabb génforrásai?
4. Milyen nemesítési módszereket használhatunk a diónemesítésben?
5. Melyek a honosítás veszélyei és nemzetközi tapasztalatai?
6. Mi a jelentősége az interspecifikus hibrideknek?
7. Melyek a legfontosabb fajtairányzatok a világ nagy diótermesztő országaiban?
8. Melyek a jelentősebb diótermesztő európai országok nemesítési és fajtahasználati irányvonalai?
9. Melyek az ideális diófajta kritériumai?
10. Melyek a magyar diópopuláció legfontosabb, nemesítésben is használható sajátosságai?
11. Mutassa be az ukrán diófajták legfontosabb sajátosságait és fajtáit!
12. Mutassa be a szlovén diófajták legfontosabb sajátosságait és fajtáit!
13. Mutassa be a francia diófajták legfontosabb sajátosságait és fajtáit!
14. Mutassa be a szerb diófajták legfontosabb sajátosságait és fajtáit!
15. Mutassa be a bolgár és amerikai diófajták legfontosabb sajátosságait és fajtáit!
16. Mutassa be a román diófajták legfontosabb sajátosságait és fajtáit!
17. Milyen szempontokat alapján zárható ki egy külföldi nemesítésű diófajta a magyarországi termesztésből?
Gesztenye
1. Milyen nemesítési módszerekkel állíthatunk elő gesztenyefajtákat? Értékelje az egyes nemesítési módszerek sikerességét!
2. Milyen nemesítési módszereket használtak a magyar gesztenyenemesítésben?
3. Milyen gesztenyefajok használhatók a gesztenyenemesítésben? Értékelje őket!
4. Milyen nemesítési célokat kell figyelembe venni a gesztenyenemesítésben?
5. Ismertesse a jelentősebb gesztenyetermesztő országok fajtairányzatait!
6. Ismertesse a magyar gesztenyefajta-használatot!
7. Ismertesse és a legfontosabb pomológiai sajátosságaikkal együtt jellemezze a külföldön nemesített, de vélhetően a magyar ökológiai adottságok mellett is ígéretesen termeszthető gesztenyefajtákat!
Mogyoró
1. Ismertesse a mogyoró termesztésének rövid történetét!
2. Melyek a fontosabb mogyorónemesítési célok?
3. Milyen génforrásokat használnak a mogyorónemesítéssel foglakozó kutatók?
4. Melyek a fontosabb mogyorót termesztő országok?
5. Milyen fajták használata terjedt el a fontosabb mogyorótermesztő országokban?
6. Melyek a jelentősebb fajtairányzatok mogyorótermesztésben?
7. Ismertesse a magyar fajtahasználat alakulását!
8. Nevezze meg és röviden jellemezze a magyar nemesítésű mogyorófajtákat!
9. Melyek a jelentősebb, ígéretes külföldi nemesítésű mogyorófajták?
A fejezet megírásához felhasznált és tanulmányozásra ajánlott irodalom:
Dió
Botu M., Tudor M., Papachatzis A. 2010/a. Evaluation of some walnut cultivars with different bearing habits in the ecological conditions of Oltenia – Romania. Acta Hortic. 861:119-126.
Botu M., Tudor Maria, Botu I., Cosmulescu Sina, Papachatzis A. 2010/b. Evaluation of walnut cultivars in the conditions of the Oltenia’s hill area regarding functioning potential. Annals of the University of Craiova, the series Biology, Horticulture, Food Produce Processing Technology, Environment Engineering, XV( XLXI ): 94-103.
Bujdosó G. (2003): Melyik a legjobb áruértékű fajta? Kertészet és szőlészet. 23(52):17p.
Cerovic, S., Golosin, B., Ninic Todorovic, J., Bijelic, S., Ognjanos, V. (2010): Walnut (Juglans regia L.) selection in Serbia. Hort. Sci. (Prague). 37(1): 1-5.
Chevallier A., Bray O., Prunet J.P., Giraud M. (2009): Factors Influencing Walnut Blight Symptoms Emergence and Development. Acta Hortic. 861. 473- 478p.
Chevallier A., Laymajoux D. (2009): A Three Year Study on the Effect of Chelal® Kubig on the Reduction of Necrotic Walnut Falls. Acta Hortic. 861. 445- 448p.
Cosmulescu S., Baciu A., Botu M., Achim Gh. 2010/a. Environmental factors' influence on walnut flowering. Acta Hortic. 861:83-88.
Cosmulescu S., Botu M., Trandafir I., 2010/b. Mineral composition and physical characteristics of walnut (Juglans regia L.) cultivars originating in Romania. Selçuk Tarım ve Gıda Bilimleri Dergisi, 24(4): 33-37.
Cosmulescu Sina, Botu M., Achim Gh. 2012/c. D etermination of apomictic fruit set ratio in several Romanian walnut (Juglans regia L.) cultivars. Not Bot Hortic Agrobo, 40(1):229-233.
Dzhuvinov, V., S. Gandev, K. Koumanov and V. Arnaudov. 2010. Walnut. Biofruit, Plovdiv. (kézirat)
Escobar, M.A., Leslie, C.A., McGranahan, G.H., Dandekar, A.M. (2002): Silencing crown gall disease in walnut (Juglans regia L.) Plant Sci 163:591-597
Frutos D., López G. (2012): Search for Juglans regia genotypes resistant/tolereant to Xanthomonos arboricola pv. juglandis in the framework of Cost Action 873. Journal of Plant Pathology. 1 (94): 1.37-1.46 p.
Gauthier M.M., Jacobs D. F. (2011): Walnut (Juglans spp.) ecophysiology in response to environmental stresses and potential acclimation to climate change. Crop Biotech Update. http://www.isaaa.org/kc/cropbiotechupdate/article/default.asp?ID=8845
Giraud M., Prunet J.P., Chevallier A., Ramain S., Thiriaud V., Santrac I., Bray O. (2009): Study of Xanthomonas arboricola pv. juglandis Population Dynamics in French Walnut Orchards over Three Years. Acta Hortic. 861. 439-443p.
Hasey J.K., Anderson K.K., Caprile J.L., Kluepfel D.A., Lampinen B.D., Grant J.A., Beede R.H. (2009): Crown Gall Incidence: Seedling Paradox Walnut Rootstock versus Own Rooted English Walnut Trees. Acta Hortic. 861. 453-455.
Hendricks, E. Lonnie, Coates, W. William, Elkins, B. Rachel, McGranahan, H. Gale, Phillips, A. Herb, Ramos, E, David, Reil, O. Wilbur, Snyder, G. Roland (1998): Selection of varieties In. Ramos, E. David (szerk.): Walnut production manual. University of California, Oakland, 84-89.
Jenkins T.A., Marsh C., Lang M.D., Vanestre J., Walter M., Obanor F. (2009): Walnut Blight Sustainable Management Research in New Zealand. Acta Hortic. 861. 479- 487p.
KSH (2003): Gyümölcsültetvény-gazdálkodás Magyarországon, 2001. Adatgyűjtemény 1. kötet. 56-59. p.
Korac M., Slovic D., Rudic M., Cerovic S., Golosin B. (1986): Rezultati hibridizacije i selekcije oraha na Poljoprivrednom fakultetu u Novom Sadu. Proceedings of the Yugoslav Symposium on the Selection and Breeding of Fruit, Cacak. 10-107p.
Leslie C., McGranahan G. (1998): The origin of the walnut, pp. 3–7. In: Ramos, D.E. (szerk.), Walnut Production Manual. University of California. Division of Agriculture and Natural Resources. Publication 3373. 319 pp.
Leslie, C.A., McGranahan, G.H., Dandekar, A.M., Uratsu, S.L., Vail, P.V., Tebbets, J.S. (2001): Development and field testing of walnuts expressing the crylA(c) gene for Lepidopteran insect resistance. Acta Hortic. 544:195-199.
Lovera M., Arquero O., Serrano N., Trapero A., (2008): Walnut blight (Xanthomonas arboricola pv. juglandis): Factors that influence the disease. Cost 873, WG and Management Committee Meeting, Athens, Greece. http://www.cost873.ch/uploads/_files/m_Athens_Abstracts_FinalBook.pdf
McGranahan, G.H., Leslie, C.A., Woeste, K.E. (1997): Backcrossing breeding walnuts for resistance to the cherry leafroll virus. Acta Hortic. 442: 121-127.
McGranahan, G.H., and Leslie, C.A. (2004).‘Robert Livermore’, a Persian walnut cultivar with a red seedcoat. HortScience 39:1772.
McGranahan G.H., Leslie C.A. (2009): Breeding of walnuts (Juglans regia) in, P.M. Priyadarshan (szerk.), Breeding Plantation Tree Crops: Temperate Species. Springer Science Business Media 249-272. p.
McKenna, J.R., Epstein, L. (2003): Susceptibility of Juglans species and interspecific hybrids to Agrobacterium tubefaciens. HortScience. 3(38): 435-439
Mohácsy M. és Porpáczy A (1951): Dió, mandula, mogyoró, gesztenye. Mezőgazdasági Kiadó, Budapest. 25. p.
Özaktan H., Erdal M., Akkopru A., Aslan E. (2007). Evaluation of susceptibility of some walnut cultivars to Xanthomonas arboricola pv.juglandis by immature nut test. Cost 873, WG3/WG4 Joint Meeting, Murcia, Spain. http://www.cost873.ch/_uploads/_files/m_ozaktan_murcia.pdf
Özaktan H., Erdal M., Akkopru A., Aslan E., (2008). Evaluation of susceptibility of some walnut cultivars to Xanthomonas arboricola pv. juglandis by immature nut test Cost873, WG and Management Committee Meeting, Athens, Greece. http://www.cost873.ch/_uploads/_files/Ozaktan_WalnutSusceptibility.pdf
Parveaud C.E., Ramain S., Verhaeghe A. (2009): The Relationship between Walnut Blight Expression and Tree Water Status. Acta Hortic. 861. 435-437p.
Ramos D.E. (Ed.) (1998). Walnut Production Manual. University of California. Division of Agriculture and Natural Resources. Publication 3373. 319 pp.
Rovira M., Moragrega C., Aletà N. (2007). Susceptibility study to Xanthomonas arboricola pv. juglandis, of immature fruits in two ‘full-sib’ walnut progenies. Cost 873, WG3/WG4 Joint Meeting, Murcia, Spain. http://www.cost873.ch/_uploads/_files/m_rovira_murcia.pdf
Ruiz-García L., Frutos D., López G., Fuentes A. (2009). Molecular characterization of walnut and evaluation of Xanthomonas arboricola pv. juglandis damages in Murcia, Spain. Annual Cost 873 Meeting, Cetara, Italy. http://www.cost873.ch/_uploads/_files/DFrutos_WalnutTyping_Italy.pdf
Solar A., Stampar F. (2005): Evaluation of some perspective walnut genotypes in Slovenia. Acta Hortic 705. 131-136.
Solar A., Jakopic J., Mikulic-Petkovsˇek M., Veberic R., Dreo T., Zadravec P., Sˇtampar F. (2008). Validation of polyphenol contents as biochemical markers for walnut blight (Xanthomonas arboricola pv. juglandis) resistance. Cost 873, WG and Management Committee Meeting, Athens, Greece. http://www.cost873.ch/_uploads/_files/m_Athens_Abstracts_FinalBook.pdf
Solar A., Dreo T., Miculic-Potrovsej M., Lizocar A., Sustarsic M., Veberic R., Maticic L., Ravnicar M., Stampar F. (2009). Phenolic compounds as potential markers for walnut blight resistance. Annual Cost 873 Meeting, Cetara, Italy. http://www.cost873.ch//_uploads/_files/ASolar_Walnut-Phenolics_Italy.pdf
Süle S., Pinter Cs., Kadlicsko S., Fischi G. (2007). Bacterial diseases of walnut in Hungary. Cost 873, WG1/WG2 Joint Meeting, Angers, France. http://www.cost873.ch/_uploads/_files/m_Sule_angers_1.pdf
Szentiványi P. (1998): Dió In: Soltész M. (szerk.): Gyümölcsfajta-ismeret és –használat. Mezőgazda Kiadó, Budapest. 336. p.
Szentiványi P (2006): Diónemesítés és fajtakutatás. In. Szentiványi P. – Kállay T.-né (szerk.): Dió. Mezőgazda kiadó, Budapest. 60-65. p.
Terpó (szerk.) (1976):Juglandaceae. Növényrendszertan az ökonómbotanika alapjaival. II. vol. 472. p.
Thiesz R., Bandi A., Tóth M., Balog A. (2007): Epidemiological survey of Xanthomonas juglandis and Gnomonia leptostyla on natural population of walnut (Juglans regia) in eastern Transylvania. International Journal of Horticultural Sciences. 4(13): 7-9p.
Tian J., Wu Y., Wang Y., Han F. (2009): Development and prospects of the walnut industry in China. Acta Hortic. 861. 31-38. p.
Tsiantos J., Vagelas I.K., Rumbos C.I., Chatzaki A., Rouskas D., Gravanis F.T. (2008). Evaluation of resistance of cultivated walnut varieties and selections to Xanthomonas arboricola pv. .juglandis in Greece. Cost 873, WG and Management Committee Meeting, Athens, Greece. http://www.cost873.ch/_uploads/_files/Tsiantos_WalnutResistanceInGreekVarieties.pdf
Vahdati K., McKenna J.R., Dandekar A.M., Leslie C.A., Uratsu S.L., Hackett W.P., Negri P., McGranahan G.H. (2002). Rooting and other characteristics of a transgenic walnut hybrid (Juglans hindsii and J. regia) rootstock expressing rolABC. J. Amer. Soc. Hort. Sci. 127:724–728.
Verhaeghe, A. (2011): France walnut cultivars (kézirat)
Wu G.L., Meng H.J., Hao Y.Y., Liu Q.L., Wang D., Tian J.B. (2009): Thirty years of breeding walnut in China. Acta Hortic. 861. 109-118. p.
Zatokovy F. T. (2009): Ukrainian cultivars of walnut. (kézirat).
Zohary D., Hopf M. (1993). Domestication of Plants in the Old World. Clarendon Press.Oxford. 249 pp.
Felhasznált internetes források:
Internet 1: http://www.walnuttrees.co.uk/shop/walnut-fruit-trees/fernor
Internet 2: http://www.coopenoix.com/enus/lacoop%c3%a9rative/lesproduits/noixfra%c3%aeches.aspx (2012. július 28-i állapot)
Internet 3: http://sacramentopacking.com/uploads/Chandler_Walnuts.JPG (2012. szeptember 3-i állapot)
{kind=link}
Gesztenye
Anagnostakis, S., Clark, S., McNab, H. (2009): Preliminary report ont eh segregation of resistance in chestnuts to infestation by oriental chestnut gall wasp. Acta Hortic. 815:33-36.
Bolvanský, M., Kobza, M., Juhásová, G., Adamčíková, K., Ostrovský, R. (2009): Spread of chestnut blight in a plantation of young Castanea sativa and Castanea hybrids. Acta Hortic. 815:261-270.
Botu, M. (2009): Sweet chestnut situation in Romania. Acta Hortic. 866:511-515.
Bratanova-Doncheva, S., Chipev, N., Dimitrova, V. (2009): Origin, state and perspective of chestnut cultivation in Bulgaria. Acta Hortic. 866:517-525.
Çeliker, N.M. és Onoğur, E. (2009): Preliminary studies on the fungal disorders especially on ink disease causing decline of chestnut trees in Turkey. Acta Hortic. 815:227-232.
Cheng, L.L., Huang, W.G. (2009): Diversity of choroplast DNA SSRs in wild and cultivated C. mollissima: Evidence of cultivated C. mollissima origon. Acta Hortic. 866:243-249.
Diamandis, S. (2009): Sweet chestnut: from the „Kastania” of the ancient Greeks to modern days. Acta Hortic. 866:527-530.
Döken, M.T. (2009): Chestnut blight and a review of the related studies in Turkey. Acta Hortic. 815:213-220.
Fullbright, D.W., Mandujano, M., Stadt, S. (2009): Chestnut production in Michigan. Acta Hortic. 866:531-537.
Heiniger, U. és Rigling, D. (2009): Application of the Cryphonectria hypovirus (CHV-1) to control the chestnut blight, experience from Switzerland. Acta Hortic. 815:233-246.
Hozova, L., Jankovsky, L., Akkak, A., Torello Marioni, D., Botta, R. és Šmerda, J. (2009): Preliminary study of the genetic structure of a chestnut population in the Czech Rebulic based on SSR analysis. Acta Hortic. 815:43-50.
Krstin, LJ., Krajačić, M., Ćurković Perica, M., Novak-Agbaba, S. és Rigling, D. (2009): Hypovirus-infected strains of the fungus Cryphonactria parasitica in the Central part of Croatia. Acta Hortic. 815:283-288
KSH (2003): Gyümölcsültetvény-gazdálkodás Magyarországon, 2001. Adatgyűjtemény, 1. kötet. 64-67. p.
Lushaj, B.M., Tabaku, V. (2009): Conersion of old, abandoned chestnut forest into simple coppice and from simple coppice forest into Orchards in Tropoja, Albania. Acta Hortic. 866:251-258.
Pandit, A.H., Kour, A., Wani, M.S. és Mir, M.A. (2009): Genetic resources of chestnuts in Kashmir Valley. Acta Hortic. 815:51-56.
Pridnya, M.V., Pinkovsky, M.D., Romashin, A.D. (2009): Novelty position of European Chestnut forests in Russia (Western Caucasus) and perspective of their protection and sustainable development. Acta Hortic. 866:543-550.
Radócz L., Tarcali G., Egyed K. (2009): The chestnut blight fungus (Cryphonectria parasitica Murr, (Barr) in the Central-European region. Acta Hortic. 866:399-404.
Sartor, C., Botta, R., Mellano, M.G., Beccaro, G.L., Bounous, G., Torello Marinoni, D., Quacchia, A. és Alma, A. (2009): Evaluation of susceptibility to Dryocosmus kuriphilus yasumatsu (Hymenoptera: Cynipidae) in Castanea sativa Miller and in hybrid cultivars. Acta Hortic. 815:289-298.
Serdar, U., Demirsoy, H. and Demirsoy, L. (2009): Determination of superior chestnut genotypes in the Central Black sea region of Turkey. Acta Hortic. 815:37-42.
Solar, A., Stampar, F. (2009): Promising chestnut genotypes from Slovenia. Acta Hortic. 866:259-264.
Soylu, A. (2009): The development of chestnut industry of Turkey in the last half century. Acta Hortic. 866:505-510.
Soylu, A. és Mert, C. (2009): The future of chestnut breeding work in Turkey. Acta Hortic. 815:27-32.
Szentiványi P. (1998): Dió In: Soltész M. (szerk.): Gyümölcsfajta-ismeret és –használat. Mezőgazda Kiadó, Budapest. 343. p.
Takada, N., Sato, A., Sawamura, Y., Nishio, S., Saito, T. (2009): Influence of pollen on pellicle removability and nut weight of Japanese chestnut (Castanea crenata Sieb. et Zucc.) ’Porotam’. Acta Hortic. 866:239-242.
Turchetti, T. and Maresi, G. (2009): Biological control of chestnut diseases in Italy: Effectiveness of blight and ink disease management. Acta Hortic. 815:253-260.
Vidóczi H., Varga M. Szabó I. (2007): Chestnut blight and its biological control in the Sopron Hills, Hungary. Acta Silvatica & Lignaria Hungarica. 3: 199-205.p
Yamanashi, O.K., Sobierajki, G.R., Bueno, S.C.S., és Pommer, C.V. (2009): Chestnut in Brazil: Reseraches and perspectives. Acta Hortic. 866:539-541.
Felhasznált internetes források:
Internet1: http://www.flickriver.com/photos/pruffoli/5095873899/
Internet2: http://www.marronedicasteldelrio.it/marroni_igp.htm
Mogyoró
Fidigelli, C., de Salvador, F.R. (2009): World hazelnut situtation and perspectives. Acta Hortic. 845. 39-52. p.
KSH, (2003): Gyümölcsültetvény-gazdálkodás Magyarországon, 2001. Adatgyűjtemény, 1. kötet. 68-71. p.
McCluskey, R.L., Mehlenbacher, S.A., Smith, D.C., Azarenko, A. N. (2009): Advanced selection and new cultivar performance in hazelnut trials planted in 1998 and 2000 at Oregon State University. Acta Hortic. 845. 67-72. p
Miljkovic, I (2000): Croatia. Hazelnut cultivar list and some important characteristics and remaks. In. Köksal, Y. (szerk.): Inventory of hazelnut research, germplasm and references. REU Technical Series No. 56. Food and Agricultural Office of the United Nations ROME, Mediterranean Agronimic Institute of Zaragosa, Spain. http://www.fao.org/docrep/003/X4484E/x4484e11.htm#P4777_40180
Mirotadze, N. (2005): Hazelnut in Georgia. Acta Hortic 686, 29-34. p.
Molnar, T.J, Capik, J.M., Gorrreda, J.C. (2009): Response of hazelnut progenies from known resistant parents to Anisogramma anomala in New Yersey, USA. Acta Hortic. 845, 73-82. p.
Pilotti, M., Brunetti, A., Tizzani, L., Gallelli, A., Lumia, V., Gervasi, F. (2009): gene candidates for pathogen perception in Corlyus avellana. Acta Hortic. 845. 115-122. p.
Rovira, M., Ferreira, J.J., Tous, J., Ciordia, M. (2005): Hazelnut diversity in Asturias (Northern Spain). Acta Hortic. 686, 41-46. p.
Solar, A (2000): Slovenia. Hazelnut cultivar list and some important characteristics and remaks. In. Köksal, Y. (szerk.): Inventory of hazelnut research, germplasm and references. REU Technical Series No. 56. Food and Agricultural Office of the United Nations ROME, Mediterranean Agronimic Institute of Zaragosa, Spain. http://www.fao.org/docrep/003/X4484E/x4484e11.htm#P4777_40180
Slyusarchuk, V.E., Ryabokon, A.P. (2005): Ukrainian hazelnuts: Cultivars, agrotechnics, perspectives. Acta Hortic 686, 603-608. p.
Thompson, M.M, Lagerstedt, H.B, Mehlenbacher, S. A (1996): Hazelnuts. In: Janick, J., Moore, J.N. (szerk.): Fruit breeding, vol. 3, Nuts. Wiley, New York, 125–184 p.
Tomcsányi P. (1979):Gyümölcsfajtáink. Gyakorlati pomológia. Mezőgazda Kiadó, Budapest.
Xie, M., Zheng, J., Radicati, L., Me., G. (2005): Interspecific hibridization of hazelnut and performance of 5 varieties of China. Acta Hortic 686, 65-70. p.
Felhasznált internetes források:
Internet1: http://hazelnut.org.nz/variety/Barcelona.html (2013. január 17.)
Internet2: http://www.njuskalo.hr/orasasti-plodovi-sjemenke/ljesnjaci-istarski-duguljasti-ljusci-1-kilogram-26-kn-oglas-5896055 (2013 . január 17.)
Internet3: http://www.agroportal.hr/agro-baza/sortne-liste/voce/ljesnjak/istarski-okrugli/ (2013. január 20.)
Internet4: http://safenut.casaccia.enea.it/db/accessions/1175 (2013. január 20.)
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028