Populációk közötti kölcsönhatások, populáció kollektívumok, növénytársulások: szerkezet, működés és dinamika
Szerző: Höhn Mária
Az élőhelyeket benépesítő populációk, együttélő közösségek egymással szorosabb vagy lazább kapcsolatban állnak. Ezen kapcsolatok természete, erőssége és iránya attól függ, hogy az egyes populációk milyen forrásokat, milyen környezeti tényezőket használnak életben maradásukhoz, gyarapodásukhoz, szaporodásukhoz. A növényi populációk közötti kölcsönhatások is akkor válnak felismerhetővé, ha tudjuk az egyes populációk igényét, helyét és betöltött szerepét az adott élőhelyen.
Ehhez abból kell kiindulnunk, hogy mely környezeti tényezőket használja egy adott populáció és azon tényezők milyen intervallumát tudja tolerálni, vagyis mi a populáció tűrőképessége. Előző fejezetekben láttuk, hogy minden környezeti tényezőre nézve megadható a populáció tűrőképessége, azaz a toleranciája. (lásd. 6 fejezet 2. ábra)
Ha egy populációnál a ható környezeti tényezők mindegyikére (n tényező) adott választ együttesen figyelembe vesszük, akkor kapunk egy sokdimenziós tolerancia mezőt, amelyet a populáció niche-nek nevezünk.
A niche (ökológiai fülke, életsáv)
- a populáció igényéből fakadó tolerancia mező
- sokdimenziós, a ható környezeti tényezők által definiálható absztrakt tér,
- a populáció jelenlétével lefoglalja, betölti.
A niche fogalom értelmezése több ökológustól is ismert (pl. Grinnel (1917) ill. Elton (1927) megfogalmazásából), de modern ökológiai szemléletű megközelítése George E. Hutchinsontól (1957, 1978) származik (1. Ábra).
Vegyünk három környezeti paramétert, és nézzük meg milyen a populáció toleranciája (válasza) a három tényezőre nézve. Mindegyik tényezőre nézve megadjuk a tolerancia határokat és összekapcsolva ezeket egy háromdimenziós tolerancia mezőt kapunk. Újabb és újabb paramétereket véve negyedik, ötödik stb. niche-tengelyekkel bővül a jellemzésünk így négy, öt stb. dimenziós niche-tereket kapunk (2. Ábra).
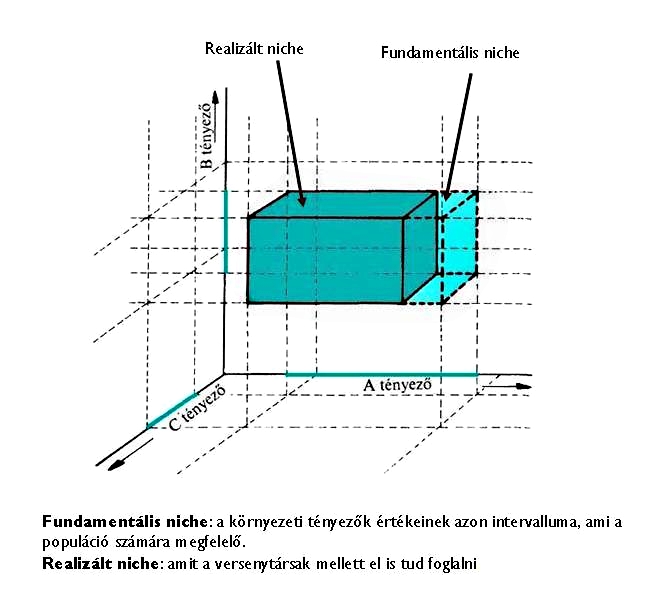
1. Ábra: Hutchinson-féle niche ábrázolás: egy 3 dimenziós tolerancia mező (lásd. Turcsányi (1998)
Megkülönböztetjük a fundamentális niche-t, mely a populáció igényéből fakadó tolerancia mező, és a realizált niche-t, melyet a populáció a versenytársak mellett el is tud foglalni. Hiszen az élőhelyen ugyanazon környezeti paraméterek akár több populáció számára is megfelelők lehetnek, vagyis többen is ugyanazt használják. A realizált niche következésképpen mindig kisebb, vagy legfeljebb egyenlő a fundamentális niche-el.
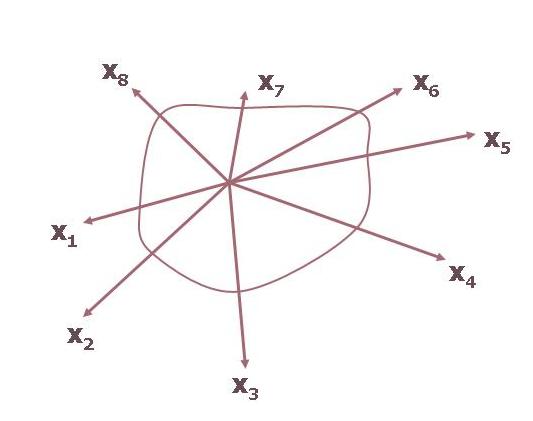
2. Ábra: A niche ábrázolása környezeti tényezők tengelyei mentén
A niche-el kapcsolatosan napjainkban számos értelmezés látott napvilágot, így például Miller szerint a niche mint a populáció „Lakcíme és foglalkozása” fogható föl. Fontos rögzíteni azonban azt is, hogy bármilyen ábrázolást és értelmezést adunk a niche nem statikus egységet fejez ki. Minden populáció niche egy „folytonos változók által kifeszített tér”. A niche-ek alapján a populációk összehasonlíthatók. Ha két populáció niche-e egybeesik, részben vagy egészben átfedődik (ún. overlap ), akkor azt jelenti, hogy a környezeti tényező(k) hasonló intervallumát használják (3. Ábra). Az átfedődő niche-ekben pedig versengés lép föl. A niche átfedés típusai:
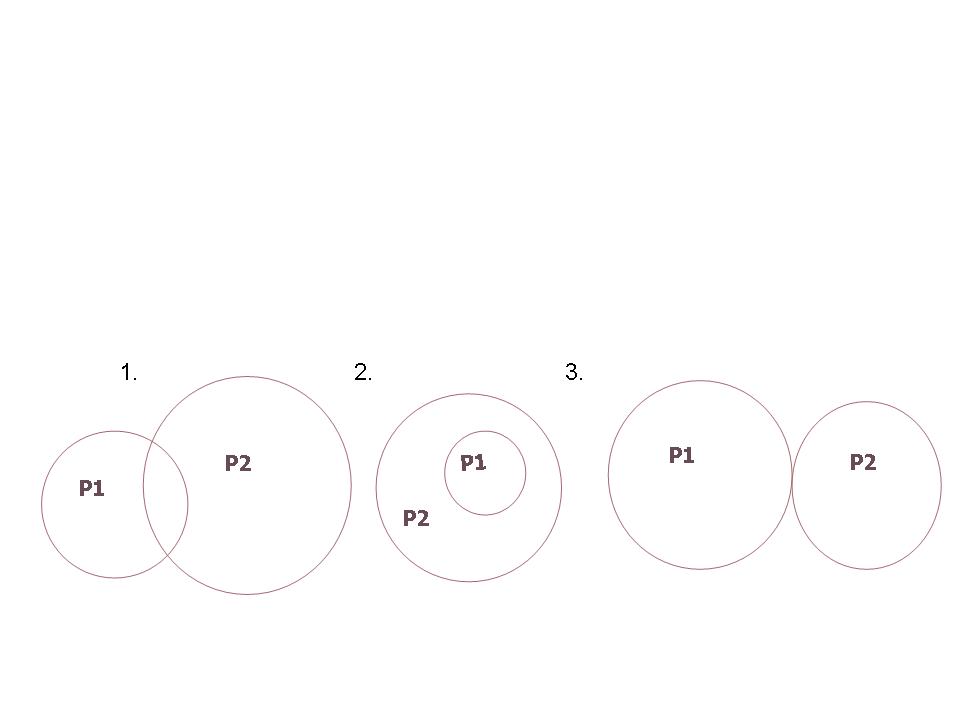
3. Ábra: A niche átfedés típusai: 1. részben átfedődő niche-k, 2. bezárt niche, 3. érintkező niche-k
A Gause-féle kizárási elv szerint, ha két populáció igényei és ezáltal niche-ük teljesen egybeesik, akkor szükségszerűen versengés indul köztük, és a kompetitív kizáródás elve érvényesül, vagyis többnyire az egyik populáció kiszorul a területről. Ha azonban a niche-k nem fednek át teljesen, vagy a kompetíció eredményeképpen szétválnak, akkor a populációk tartósan együtt maradhatnak, ún. koegzisztencia alakulhat ki. Ebből mindjárt az is következik, hogy egy élőhelyen azon populációk lesznek együtt, alkotnak közösséget, melyek niche egymás mellett vagy legalábbis egy grádiens mentén szerveződik. A niche grádienst a populációk optimumgörbéinek a niche dimenziók mentén való ábrázolása rajzolja ki (4. Ábra).
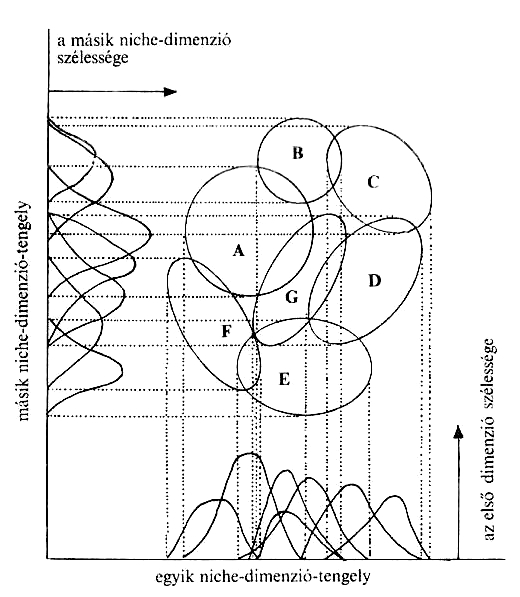
4. Ábra: Niche grádiens (átfedés és eloszlás mértéke) Széky Pál nyomán
7. 2. Populációk közötti kapcsolatok
A niche alapján értelmezhetjük a populációk közötti kapcsolatokat is. Bizonyos fajok populációi között nincs kimutatható, érzékelhető kapcsolat, ezt nevezzük neutralizmusnak, mely egymástól jól elkülönülő niche-k esetén valósulhat meg.
Ha bármilyen kapcsolat kialakul két populáció között, akkor az egymásra hatás típusa szerint beszélhetünk:
- negatív kapcsolatról, mely lehet
kölcsönhatás vagy interakció: ilyen a versengés vagy kompetíció
egyoldalú befolyásolás: ilyen a parazitizmus és az allelopátia
- pozitív kapcsolatról: mint a szimbiózis, a mikorrhiza és a mutualizmus
- laza, kisebb túlélési előnyökön alapuló kapcsolatokról: mint az asztalközösség vagy kommenzalizmus (5. Ábra)
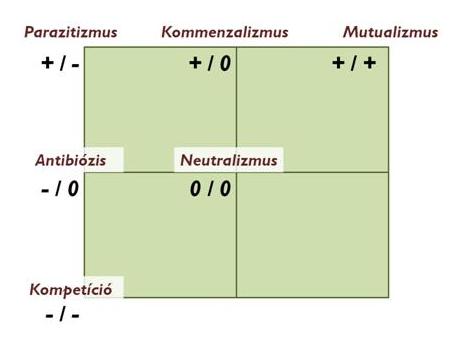
5. Ábra: A populációk kölcsönhatásának típusai illetve várható eredményei
A versengés vagy kompetíció
Az élőhelyek populáció közösségeiben leggyakrabban előforduló kapcsolat, mely nagy változást okoz a populációk életében és rendszerint az egyedszám szignifikáns csökkenésével jár. A kompetíció mindig az elfogyó környezeti tényezőkért folyik, és a legjobb, legsikeresebb versengő jut hozzá a forráshoz. Forrásnak tekinthető minden olyan elfogyó tényező, amely elengedhetetlen a populáció életbenmaradásához, fejlődéséhez, ezért a forrásokért mindig versengés alakul ki (átfedődő niche-k).
Az élőlények életében néhány forrás kitüntetett fontosságú. Az autotróf növényeknél ilyen a fény, a víz és a talajban lévő tápanyagok. Bármelyik tényező ezek közül eléri a populáció tűrőképességi határait ún. limitáló tényezőként lép fel. Kritikus értékei a populáció kipusztulását jelentheti. Nem minden elengedhetetlen környezeti tényező egyben limitáló tényező, mert például a légköri CO2, mely a fotoszintézis alapeleme, koncentráció értékeivel folyamatosan a populációk rendelkezésére áll. A Liebig–féle minimum elv a talaj tápanyagtartalom elérése vonatkozik és kimondja, hogy mindig a minimumban lévő tápanyag mennyisége határozza meg az egyedek fejlődését függetlenül attól, hogy a többi tápanyag milyen mennyiségben van.
A versengés eredménye vagy kimenetele kétféle lehet:
- A kompetitív kizáródás során egyik populáció eltűnik az élőhelyről.
- A koegzisztencia során viszont mindkét populáció kisebb egyedszám mellett, a realizált nichének megtartásával, igazításával folytatja a források felhasználását.
Az „igazítás” az élőlények, populációk viselkedéséből adódik, mennyire képesek plasztikusan kezelni a környezeti változásokat, ez pedig a niche potenciáljukból adódik.
Így a kiaknázó kompetíció esetén a versenyben lévő populációk forrás hasznosításának hatékonysága számít. Aki több forrást tud kiaknázni, ezáltal jobban és gyorsabban növekszik, vagy szaporodik, meg tudja előzni a másikat. Ennek következtében a másik versengő fél növekedési, szaporodási esélye lecsökkenhet, és emiatt további forrásfelhasználásról marad le. A föld alatt zajló gyökérkompetíció egyértelműen a kiaknázó kompetíció kategóriába esik, mert amelyik fél gyökere hosszabb és szerteágazóbb, az fogja sikeresebben, nagyobb kiterjedésben a talajforrásokat felhasználni. Minden populáció a sikeres versengés érdekében kompetitív stratégiák kidolgozására kényszerül. Ezeket gyakran kísérleti úton figyelhetjük meg.
Lágyszárú növényfajokból álló közösségekben egyik gyakori interspecifikus kompetíciós jelenség a Schmid és Harper (1985) által leírt ún. falanx és gerilla stratégia érvényesülése (6. Ábra). A tömött tőlevélrózsával rendelkező növények a falanx típusba tartoznak és a kompeticíó során azt a stratégiát alkalmazzák, hogy tőlevélrózsáikkal viszonylag gyorsan lefedik a talajfelszínt, gátolva ezzel más növényfaj megtelepedését. A falanx példanövénye a szászorszép (Bellis perennis), de hasonlóan viselkednek más tőlevélrózsás kétszikűek is, mint a Hieracium pilosella vagy a Plantago lanceolata.
A gerilla típusú fajok stratégiája azt jelenti, hogy a versengésre kényszerülő növényegyedek heverő szára (tarackja, rizómája) gyorsan nő, megnyúlik és túlnőve a kompetíciós teret új felszínen képes legyökeresedni. A gerilla típusnövénye a Prunella vulgaris, de hasonlóan viselkednek a Fragaria fajok is (pl. kerti szamóca). Felmerül a kérdés, hogy melyik stratégia a sikeresebb? A válasz az, hogy ez a kiinduló helyzettől függ, mert ha a kezdeti egyedsűrűség nagy, a gerilla jó eséllyel jut kompetíciós előnyhöz. Ha azonban egy csupasz felszín kiaknázásáért folyik a küzdelem a falanx stratégia bizonyul helyes stratégiának.
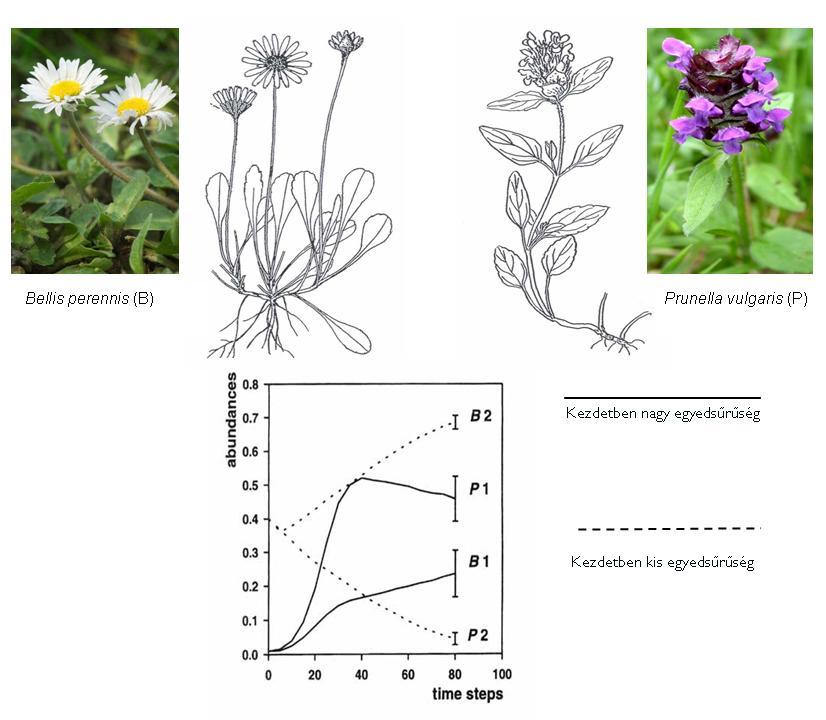
6. Ábra: A falanx és a gerilla stratégia típusnövényei és kísérletes szemléltetése (Schmid - Harper 1985 nyomán )
A kompetícióban minden „érdekelt” részt vesz. Így beszélhetünk interspecifikus (különböző fajok egyedei közötti), és intraspecifikus (fajon belüli) kompetícióról is. A fent említett példa az interspecifikus kompetíció esete.
Az intraspecifikus versengésben ugyanazon faj egyedei küzdenek és az erősebb egyed győz. A növényeknél a gyorsabban növő, nagyobb testméretű, szerteágazó gyökerű egyed vélhetőleg sikeresebb lesz. A kezdeti szakaszban megszerzett előny később kamatozódik és más források is jobban elérhetővé válnak. Gondoljunk csak a ledőlt erdő alján fejlődő magoncokra. Amelyik leghamarabb leveleket fejleszt, az gyorsabban tud nőni és beárnyékolja a társát. A fényért való sikeres küzdelem következtében az egyed növekedési erélye pedig előbb-utóbb a nagyobb gyökérzet fejlesztésében is megmutatkozik, miáltal a talajforrásokat is sikeresebben fogja kiaknázni.
Az intraspecifikus versengés az utód nemzedék fejlődésében kulcsfontosságú, hasonlóan például az egy fészekaljban lévő madárfiókákhoz. Ha az anyamadár kevés táplálékot hoz, akkor az erősebben nyújtózó, vergődő fióka veszi el a táplálékot, akár testvérétől is, aki így fokozatosan legyengülve még egy későbbi nagyobb mennyiségű táplálék esetén is elveszti esélyét a túlélésre az egyre erősebb, falánkabb testvére mellett.
A növényeknél azonban, az intraspecifikus kompetíció más formájával is találkozunk, mégpedig a modulok közötti (lényegében „egyeden belüli”) kompetícióval.
A növények testfelépítéséből adódó moduláris szerkezet, lásd Növények diverzitása fejezet, lehetővé teszi azt, hogy az egyed a modulok dinamikájával szabályozza túlélési esélyeit. A modulok közül a versengésben résztvevők ugyanúgy viselkednek, mint a populáció egyedei. Amelyik gyorsabban nő több fényhez jut, amelyik hamarabb legyökeresedik, az több tápanyagot képes felszívni. A klonális fajok moduljaik számának változtatásával, a modulok életciklusának szabályozásával sikeresebbek tudnak lenni akár az interspecifikus, akár a faj egyedei közötti, intraspecifikus kompetícióban.
A málna (Rubus idaeus) esetében például tudjuk, hogy az egyedek a föld alatti gazdag rizóma rendszerükkel képesek egy terület talajadottságait maximálisan kiaknázni. A hegyvidéki málnásokban hosszú ideig alig van más elegy faj. A málna sűrű, földalatti hajtásrendszerének rügyeiből számos modul fejlődik, melyek előbb-utóbb egymást árnyékolják, egymás növekedését, termőképességét befolyásolják (7. Ábra). A málna modulok élettartama a természetben három év. Első évben kifejlődik a zöldhajtás modul, sok levéllel. A második éves modulok virágzanak és teremnek. A harmadik éves modul már többnyire nem terem és gyenge levélzete csak a befásodott hajtások csúcsán fejlődik. Ily módon az intraspecifikus kompetíció csökkentése érdekében a modulok között a tápanyag és fénykihasználás miatt funkcionális megoszlás mutatkozik. Ez nagymértékben növeli az egyedek rátermettségét és biztosítja más fajokkal szemben a málna kompetíciós előnyét.
A málna termesztése során a termesztők is figyelembe veszik a modulok életciklusát, és mindig eltávolítják az értéktelen modulokat, ugyanakkor nem bántják az elsőéves hajtásmodult, hiszen az nem haszontalan csak a következő évben fordul majd termőre.

7. Ábra: A málna (Rubus idaeus) és klonális szerkezete
A szőlőtőkéken alkalmazott hajtásválogatás is a modulok közötti kompetíció csökkentését szolgálja. Következésképpen a moduláris testfelépítésből adódóan a növények esetében létezik az infraindividuális (egyeden belüli) kompetíció lehetősége, mely valójában a genetek szintjén a rametek zajló kompetíció.
Az allelopátia
A kompetícióban való sikeresség stratégiájának egyik sajátos formájának tekinthető az amikor az egyik partner negatív hatást gyakorol a másik populációra kémiai gátló mechanizmus révén. Bár a jelenséget régóta ismerték, a fogalom első leírója Molish (1937) volt. Egyes növények élő földfeletti vagy földalatti szerveiken keresztül, esetleg az elhalt, lehulló részeikkel olyan anyagokat, vegyületeket juttatnak a talajba, amellyel más faj egyedeinek fejlődését, csírázását gátolják.
A dió (Juglans regia) leveleinek juglon tartalmáról tudjuk, hogy allelopatikus hatású. A pillangós virágú növények levelében lévő kumarin típusú vegyületek is erős csírázásgátlók. Tapasztaljuk például azt is, hogy az akácosok gyepszintjében alig található lágyszárú növény, és az is csak kora tavasszal, amikor a talaj nedves és felhígítja az allelopatikus mérgeket (8. Ábra). A Fagales rend tagjainak cseranyagai, a tanninok szintén csírázásgátlókként működnek. Gyakran az anyanövény szabályozza, korlátozza az utódnemzedék fejlődését is, azzal hogy autotoxicitást okoz kémiai anyagok kibocsájtásával.
Sok esetben nem tudjuk az allelopátiát okozó vegyületek összetételét, de az biztos, hogy sokféle közösségben, így gyomos területeken és más száraz élőhelyi környezetben az allelopatikus hatás erős kompetíciós előnyt biztosít. A száraz élőhelyek talajában felhalmozódó allelopatikus toxinok egyre erősödő hatását a spontán tüzek szüntethetik meg. Szavannák bozóttüzei például hasznosak a vegetáció fejlődése számára, mert elégetik a felhalmozódó allelopatikus anyagokat.

8. Ábra: akácerdő tavaszi és nyári képe( forrás: http://akacosut.blogspot.hu)
Parazitizmus
Olyan egyenlőtlen kapcsolat a populációk között, melyben megváltozik a gazdanövény populációsűrűsége és a parazita létfeltételei teljesülnek.
Formái:
A. Obligát paraziták azok, akik mindig függenek a gazda közvetlen jelenlététől:
- Lehetnek teljes paraziták, amikor minden szükséges tápanyagot a gazdanövénytől vesznek fel. Ilyenek a parazita gombafajok pl. a lisztharmat gombák, a szőlő peronoszpóra (9. Ábra), vagy a szőlő szürkepenész. A virágos fajok között is találunk ilyeneket. Néhány növénycsaládban a fajok mindegyike teljes parazita, például az arankák (Cuscuta sp.), a fenyvesekben élő fenyőspárga (Monotropa hypopitys 10. Ábra), vagy a vajvirágok (Orobanche sp. 11. Ábra ). Ezeknek a virágos növényeknek egyáltalán nincsenek színanyagaik, fotoszintézisre nem képesek

9. Ábra: A nemes szőlő (Vitis vinifera) levelén a lisztharmat gomba (Plasmopara viticola) tünetei

10. Ábra: fenyőspárga (Monotropa hypopitys). Bottlik Gábor felvétele

11. Ábra: borostyán vajvirág (Orobanche hederae) Barabás Sándor felvétele.
- Félparaziták azok a fajok, amelyek csak a fotoszintézis kiinduló alapanyagait szerzik be a gazdától, de színanyagaik révén képesek fotoszintetizálni. Ilyen például a fehér fagyöngy (Viscum album) (12. Ábra), vagy a fakín (Loranthus europaeus)

12. Ábra: fehér fagyöngy (Viscum album)
B. Fakultatív paraziták olyan fajok, amelyek bizonyos körülmények között szaprofita életmódot is képesek folytatni, de elsősorban parazita életmódúak. Gombák között találunk ilyeneket.
A parazita egyed helyzetére vonatkozóan megkülönböztetünk ektoparazitákat, melyek a gazdaegyed felszínén élnek és különböző szívóképlettel hatolnak be a gazdanövénybe, valamint endoparazitákat, amelyek behatolnak a gazdaszervezetek szöveteibe. Utóbbiak között található számos gomba és baktérium faj.
Külön csoportot alkotnak a gombák között az ún. sebparaziták, ilyenek egyes taplógomba fajok, pl. Fomes, Phellinus (13. Ábra), melyek csak a gazdaszervezet sebfelületein át tudnak megtelepedni a gazdanövényen.

13. Ábra: parázstapló (Phellinus igniarius)
Különböző táplálkozási kapcsolatok, a herbivória és következményei
Nitrogénben szegény élőhelyeken így tőzeglápokban, trópusi sovány talajokon rovarfogó növények élnek. Ezek a nitrogénforrás elérése céljából különböző csapdákat állítanak rovarok számára. A csapdába került rovar vagy más éppen más kisebb ízeltlábú testét a növény emésztő válladékok révén lebontja és ezáltal nitrogén utánpótláshoz jut (14. Ábra).

14. Ábra: kancsóka (Nepenthes sp.) A, harmatfű (Drosera sp.) B, C fajok (Solymossy G. felvétele ).
A növények és a növényevő állatok között kialakuló táplálkozási kapcsolat, az ún. herbivória következtében a növényi populációk egyedszáma csökken és a növényi egyedben sérülések, sebek, csonkulások keletkeznek. Elmaradhat a virágzás és erőteljesen lecsökkenhet maghozam is A túllegeltetés nyomai legtöbbször jól megfigyelhetők (15. Ábra). A növényi egyedek védekezésképpen gyakran toxinokat termelnek, szúrós képleteket fejlesztenek, nehezen fogyaszthatóvá próbálnak válni. A rovarrágással szemben védelmet nyújthat az erős szőrözöttség vagy a fejlett kutikula is. A túllegeltetés nyomait gyakran könnyű felismerni, mert némelyik mérgező növényfaj nagyon felszaporodik, és közötte számos szúrós növény is megjelenik. A sok szilárdító szövettel rendelkező és vízhiányos leveleket kevesebb állat fogyasztja.

15. Ábra: A túllegeltetés nyomai egy délies kitettségű lejtőssztyepben (bal) és egy szikes pusztán (jobb)
A kölcsönös előnyökön alapuló ún. mutualisztikus kapcsolatok
A szimbiózis és mikorrhiza
A kölcsönös előnyökön alapuló populációk közötti kapcsolatok közül sok azért jött létre, hogy a partner populációk ezáltal sikeresebbek legyenek a más fajokkal fennálló kompetícióban, illetve jobban hozzáférjenek a forrásokhoz. A pozítiv jellegű populációk közötti kapcsolatoknak fontos szerepe van a szélsőséges élőhelyek benépesítésénél és a nagy fajsűrűségű élőhelyeken való életben maradásnál.
A növényvilág evolúciójában a hajtásos növényfajok között alakult ki a nitrogén fixációra szolgáló szimbiózis képessége és ezen fejlődési vonalakhoz tartozó taxonok nagy kompetíciós előnyhöz jutottak. A talaj nitrogénhiánya ugyanis nagymértékben korlátozza a növények fejlődését, elterjedését. Minden olyan populációk közötti kapcsolat, amely segített a légköri nitrogén hasznosításában, egyértelmű kompetíciós előnnyel járt. A pillangós virágú növények (Fabaceae család) fajai a gyökereiken élő baktériumok segítségével képesek a légköri nitrogént hasznosítani. Ezek a gyökér gümőkben élő Rhizobium baktériumok számukra felvehető formájú nitrogént „gyártanak” (16. Ábra).
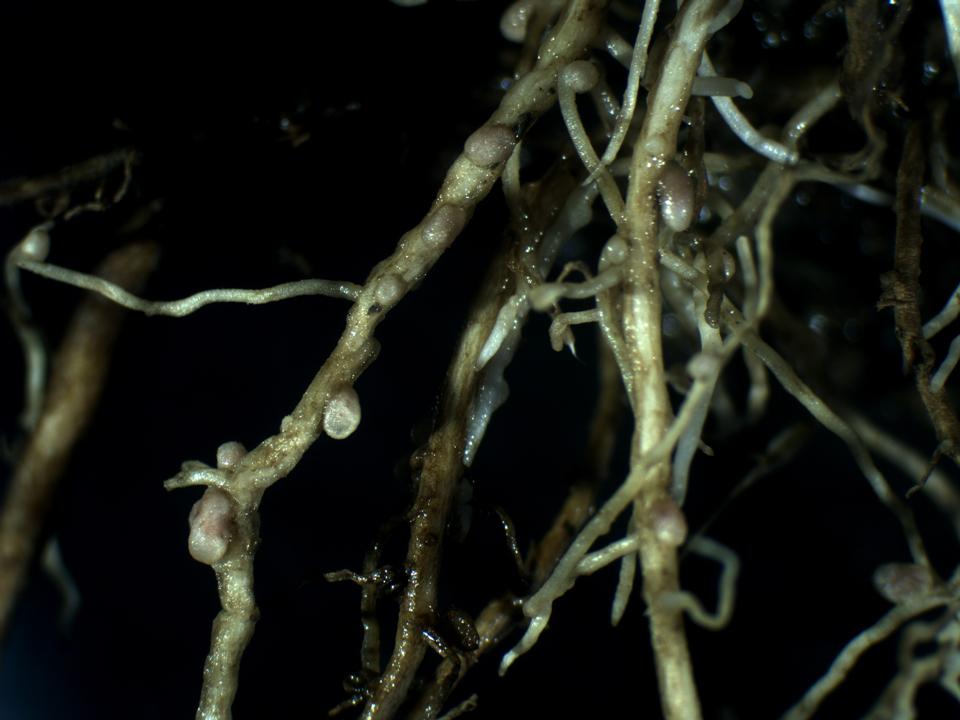
16. Ábra: gyökérgümős gyökér a pillangós virágúaknál.
Ismert szimbiotikus kapcsolat alakul ki a már korábban említett Actinomycetales és az éger fajok között. Az éger (Alnus) fajok a szervetlen nitrogénben szegény talajokon nőnek ezért elengedhetetlen számukra a nitrogén felvétel segítése.
A gomba-alga együttélés egy másik evolúciós sikerhez vezetett a zuzmók megjelenése által. A zuzmó nem gomba és nem alga, hanem egy fejlődéstörténetileg új csoport, mely a sikeres együttélés, szimbiózis eredményeképpen jött létre. Sem a gomba, sem az alga nem képes például a zuzmósavat előállítani, tehát ez a vegyület már az új típusú szerveződés (élőlény) anyagcseretermékének számít. Ebben a sikeres együttélésben a gomba számára az alga tápanyagot állít elő autotróf módon, míg az algasejtnek kiváló védelmet nyújt a gomba szövedék. Egyes vélemények szerint valójában a gombafonalak csapdájába esett moszatot a gomba táplálék forrásként használja. Ilyen értelemben ez a jelenség egyfajta parazita kapcsolat felé mutat.
A zuzmók olyan sikeres élőlények, hogy a legcsupaszabb sziklafelszíneken és falakon is képesek megélni, illetve olyan extrém élőhelyeken, a sarkvidéktől a trópusokig, ahol önmagában sem a gomba sem az alga nem lenne életképes (17. Ábra).

17. Ábra: rénszarvas zuzmó (Cladonia sp.) és sárga falizuzmó (Xanthoria parietina)
A környezeti források jobb eléréséért a magasabbrendű növények gombafajokkal is létesítenek mutualisztikus kapcsolatot. Ilyen például a hajtásos növények gyökere és a gombafonalak együtteseként kialakuló mikorrhiza-kapcsolat. A gombafonalak sűrű szövedéke megnöveli a felszívási felületet és segíti a tápanyagszegény talajokban élő növények víz- és anyagfelszívását. Egyes mikorrhiza kapcsolatok a gyökereket megvédik a patogénhatásoktól és ezáltal a mikorrhizás egyed életképesebb lesz. A gombafajoknak cserébe a növény szénhidrátokat ad át, vitaminokkal, hormonokkal és egyéb növekedés regulátorokkal segíti a gomba fejlődését, termőtest-képzését. A hajtásos növényfajok több, mint 90%-a rendelkezik valamilyen mikorrhiza partnerrel.
A mikorrhiza kapcsolatok csoportosíthatók aszerint, hogy hol helyezkednek el a gombafonalak, így beszélhetünk ekto- és endomikorrhizáról. Az ektomikorrhizánál a gombafonalak a gyökerek felszínén alkotnak sűrű szövedéket. Illetve csak a legfelső kéregrétegekig hatolnak be a gazdanövény sejtjeibe. Az endomikorrhiza esetében viszont a gombafonalak behatolnak a sejtek belsejébe, ahol különböző képleteket hoznak létre.
Aszerint, hogy a gombák által létrehozott képletek milyen típusúak többféle csoport különíthető el (18. Ábra):
- ektomikorrhiza esetén a gyökér felületén egy gombafonál köpeny képződik és az intercellulárisokba behatoló gombafonalak az ún. Hartig-hálót alakítják ki. Főképpen fás fajokra jellemző mikorrhiza típus, sok nyitvatermőnél és lombos fánál fordul elő. A gomba kalapos, bazidiumos illetve aszkusos gomba lehet. Erdeinkben élő nagygombáink többsége mikorrhiza kapcsolatban él.
- vezikulo-arbuszkuláris mikorrhiza (VAM), amikor a kéregsejtekbe és az intercellulárisokba behatoló gombafonalak hólyagszerű képleteket ún. vezikulumokat, hurkokat illetve kisebb fákra emlékeztető hausztóriumokat alakítanak ki. Bolygatott élőhelyek fajai alkotnak ilyen mikorrhizát. Tehát a gomba partner csak a növényi sejtek belsejében képes megélni.
- erikoid mikorrhiza a hangafélék családjában alakult ki (Ericaceae), endomikorrhiza kapcsolat mely során a gomba fonalak a hajszálgyökerek sejtjeibe hatolnak be, és ott létrehoznak gombolyagszerű képleteket. A savanyú talajú területeken élő hangák nitrogén és foszfor felvételét segítik ezek a szaprofita életmódot folytató gombafajok.
- orchidea típusú mikorrhiza az Orchidaceae családra jellemző és segíti a tápanyagnélküli magvak csírázást azok kezdeti szakaszán. A későbbiekben a gomba-növény kapcsolata a parazitizmus kialakulásáig is elvezethet.
A mikorrhiza leírását a lásd. a Növényi szaporodásbiológia fejezetben.
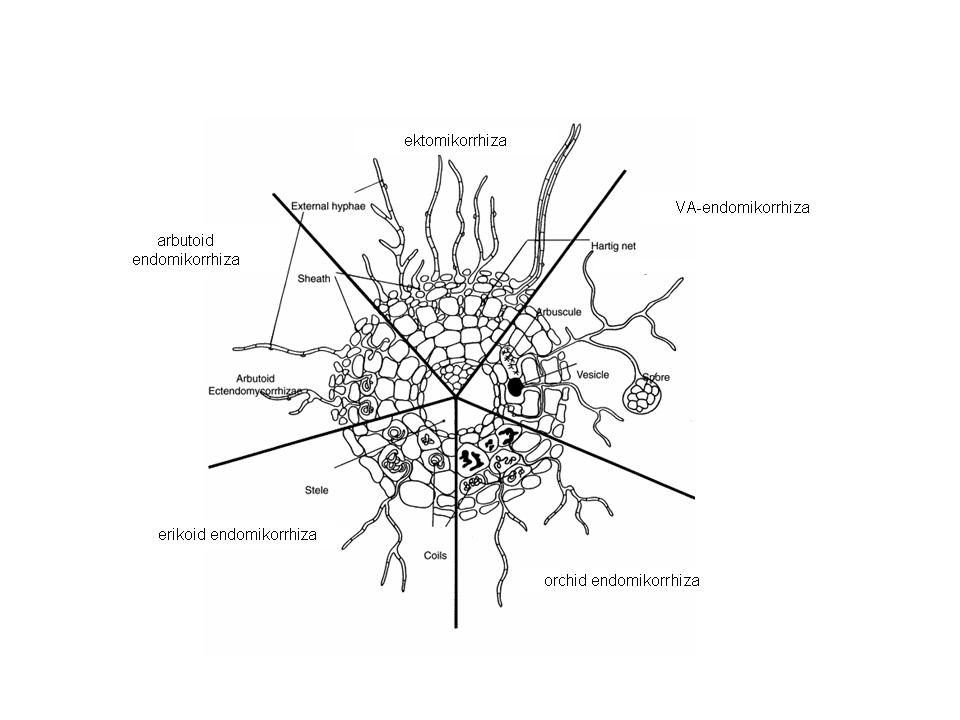
18. Ábra: Négyféle mikorrhiza típus vázlatosan
Mutualisztikus kapcsolatok léteznek más célból is a virágos növények körében. Az ún. megporzási mutualizmusok során a földi élet elsődleges szerves anyagot előállító növényei szolgáltatásaikért cserébe az állatok segítségét veszik igénybe.
Jól ismert az orchideák és megporzóik közötti szoros kapcsolat. A növény nem bízza a véletlenre a megporzását és irányítottan helyezi pollenjét egy terjesztő partner, a megporzó gondjaira. Az állatfaj számára a pollen és a nektár táplálékforrás. Ezért, ha a növényt megtalálja, azáltal hogy a virág külső jegyeiben felismerhetővé válik számára, biztos táplálékforráshoz jut. A bangófélék (Ophrys) a hím rovarok nőstények iránti vonzalmát használják ki, akik mézajkukat a megporzó rovar nőstényének képére alakítják ki (19. Ábra). A trópusokon a sokporzójú mimozafélék rendszerint denevérmegporzásúak, mert a denevérek eleségképpen fogyaszjták a porzókat miközben elvégzik a megporzást is.

19. Ábra: Ophrys arachnitiformis (pókképű bangó)
Néha a növények kegyetlenebb eszközök használatától sem riadnak vissza, növényi csapdákat állítanak fel a rovarok számára. Ilyenek például az Aristolochia nemzetség fajai, ahol a virág morfológiája úgy alakul, hogy a pollenhozó rovart csapdába ejti.
Kommenzalizmus
Olyan populációs kölcsönhatás típus, melyben az egyik populációra (gazda) gyakorolt hatás nem jelentős (nincs), de a kommenzalista populáció számára a gazda jelenléte pozitív hatású. Régóta ismert tény, hogy a fákra kapaszkodó liánok számára a fa fontos támaszték, mely által több fényhez juthat. Ha a lián nem terjeszkedik túlságosan (térparazita) akkor a fás növény ezt jól viseli, rá nézve ez nem jelent befolyást. Ugyanakkor egy tüskés, szúrós lián akár meg is védheti a fa kérgét is az arra járó rágcsáló állatoktól.
Kertekben, kisebb parcellákban gyakran látni, a kommenzalizmus gyakorlati alkalmazását is. A kukorica tövénél növő tök az erős besugárzást nehezen viseli, őt a kukorica lombja árnyékolja. Ugyanakkor az is igaz, hogy nagy széles levelei képesek beteríteni a talajt, meggátolva annak gyors kiszáradását. A kukorica, amiért árnyékot nyújt a töknek cserébe nedvesebb üdébb talajon növekedhet. A kommenzalista kapcsolatok igen sokfélék, gyakran állatok és növények, állatok és gombák populációit tartják tartósan együtt az élőhelyeken.
A populációs kölcsönhatások evolúciós következményei
A szoros együttélés gyakran evolúciós lépést eredményez, hiszen például a megváltozó virágfelépítés hatására a megporzó állatfaj populációja táplálkozási szokásait változtatja meg, mely révén elkülönül, elszigetelődik a faj többi populációjától, önálló ökotípust alakítva ki. Az új ökotípus elterjedésével új alfaj illetve hosszútávon új faj is létrejöhet. Néha nem is tudjuk, hogy melyik mutualisztikus partner evolúciója volt előbb. Ezt az együtt fejlődést koevolúciónak hívjuk.
A Ficus fajok, így a mediterránban honos füge (Ficus carica) jellegzetes serleg virágzatát a füge darázs porozza be. A füge zárt serlegvirágzatán egy bejárat van, amelyet a fügedarázs jól ismer. Egyikük sem képes szaporodni a másik nélkül, hiszen a darázs hozza a pollent a füge virágzatra, míg petéit a füge húsos (tápanyagdús) serlegvirágzatának belsejébe helyezi. A füge gyümölcs ily módon táplálékforrást és védelmet nyújt lárvái számára.
Kertészek jól tudják, hogy nálunk a mediterrán füge termései nem csíraképesek, mivel rendszeresen elmarad a megporzás. Sem a fügedarázs sem a füge elterjedési területe nem ér el hozzánk, így a termeszett füge nem porzódik be.
A védekezés, életbenmaradás érdekében néhány különleges jelenség is létrejöhetett, mint az állatok mind a növények körében.
A mimikri jelensége azt jelenti, hogy egyes fajok rejtőzködés céljából erősen hasonlítani próbálnak más populációk egyedeire vagy a környezet elemeibe igyekeznek beolvadni, láthatatlanná válva. Leginkább látványos a botsáskák mimikrije (20. Ábra), de a sarkvidék nyílt élőhelyeinek állatai is kiváló mimikrit alkalmaznak.

20. Ábra: Bengáli botsáska (Medauroidea extradentata) (Simig Dániel felvétele)
A kultúrnövények gyomközösségeiben is gyakorta előfordul, hogy a gyomnövény hajtását terméseinek formáját, színét a kultúrnövény képére igyekszik formálni. Ilyen eset például a gomborka (Camelina) és a termesztett len (Linum usitatissimum) közötti együtt-előfordulás kapcsolata (21. Ábra). Egyes források szerint a gomborka (Camelina sativa) valamikor olajáért termesztett növény volt és nem lehetett ritka, hogy lennel együtt ültették vagy éppen termesztették. Az is lehet, hogy az emberek nem irtották ki, mint gyomnövényt a lentáblából haszna miatt. Az együttes fejlődést az ember is segíthette és eredményeképpen alakulhatott ki ez a fajta idomulás a két faj között.
HIÁNYZIK
21. Ábra: foto egy gomborkás len ültetvényről .
Növénytársulások és társulásdinamika
A niche-elmélet szerint az élőhelyeken azok a populációk fordulhatnak elő, amelyek számára a környezeti tényezők paramétereinek alkalmas intervalluma található meg, vagyis minden tényező a populáció toleranciahatárain belül van. Természetesen, egy élőhely mindig több populáció számára is alkalmas lehet, így a források eléréséért ádáz küzdelem folyik. A küzdelem, amely a populációk közötti kapcsolatok révén szabályozódik, végül elvezet egy olyan állapothoz, amelyben egy olyan populáció-közösség jön létre, ahol a niche-ek nem vagy már alig fedődnek át és így a populációk tartós együttélésre rendezkednek be.
Minden élőhelyen viszont igyekszik minél több populáció fennmaradni, ezért a kompetíció eredményeképpen nichetér-felosztás következik be. A niche-tér felosztása vagy niche-szegregáció több dimenziót érinthet. Idő faktor esetén a niche szegregáció lehet szezonális és diurnus.
Szezonális niche tér felosztás mutatkozik lombos erdeink aljövényzetében is. Tavasszal, amikor még sok fény jut a talaj felszínére fényigényes lágyszárú fajok népesítik be az erdőt. Ezek jobban viselik viszont a hidegebb talajt is. Később, ha már a lombkorona zárul, ezek a fajok eltűnnek (hagymák, rizómák és gumókba visszahúzódva várják a következő tavaszt) és átadják helyüket a melegigényes, de árnyékot jól viselő fajoknak. A niche tér ilyen típusú felosztása az erdők aszpektus váltását eredményezi. A kora tavaszi aszpektus általában színesebb és virágosabb, hiszen a több fény és a tavaszi üde talaj sok növényfajnak ideális környezetet teremt (22. Ábra).

22. Ábra: gyertyános-tölgyes erdő nyári (fent) és koratavaszi (lent) aszpektusa (fent Bölöni János felvétele)
A niche tér felosztása napszakok szerint is törtéhet. Ez a jelenség inkább a mozgó állatok között figyelhető meg. Talán mindenki tudja, hogy azon a parlagon, réten ahol nappal egerészölyv vadászik éjszaka a füles bagoly keresi zsákmányát. A niche szegregáció következtében egy élőhely nagyon sok populációt képes eltartani, de csak azokat, amelyek toleranciája egybeesik a környezeti paraméterekkel. Sok populáció igyekszik ugyanakkor a niche-t az élőhelyhez „igazítani”, és ezért adaptív jellegek folyamatos kialakulása figyelhető meg. Azt mondjuk, hogy egy megtelepedő populáció valójában folyamatosan idomul a környezetéhez toleranciájának igazításával, és ezek a finom összehangolódások végülis az ökológiai adaptációhoz vezetnek el.
Egy élőhely populáció közössége akkor kerül egyensúlyi helyzetbe, ha a kompetíciós hatások csillapodásával és a pozítív kölcsönhatások erősödésével a niche-ek eltolódása végbemegy és niche-gradiensek alakulnak ki. Egy beállt növénytársulás populációinak sohasem azonos a niche-e. A niche-tér finomfelosztása miatt az élőhelyen a populációközösségek társulásokat, ún. cönozisokat építenek fel. Mivel a növények helyhez kötöttek, társulásaik állandó megjelenésűek és az élőhely mintázatát alakítják ki (23. 24. Ábra).
A növénytársulás - asszociáció - tehát:
- törvényszerűen ismétlődő, állandó megjelenésű és faji összetételű,
- meghatározott struktúrájú növényközösség,
- hasonló élőhelyi körülmények között mindig kialakul.

23. Ábra: Középhegységi szubmontán bükkös a Magas- Bakonyban
Ökológiai tényezők Társulások
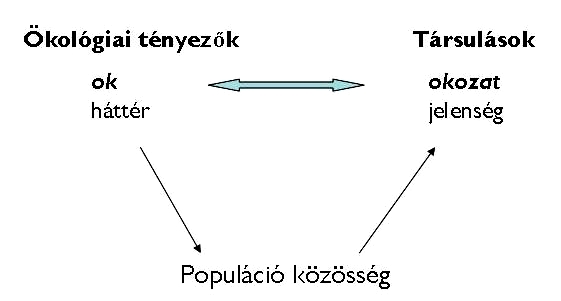
24. Ábra: A társulások kialakulásnak vázlatos ábrája
Az asszociáció (növénytársulás) a vegetáció alapegysége:
Törvényszerűen ismétlődő, állandó megjelenésű és faji összetételű, meghatározott struktúrájú növény együttes, melynek állományai hasonló környezeti feltételek között alakulnak ki. Jellemző fajok populációi alkotják, melyeket - karakter fajoknak – nevezünk. Középhegységeinkben például, 600m tengeszint feletti magasságtól bükkösök alakultak. A szubmontán és montán bükkös növénytársulások fássázárú karakterfaja a középeurópai elterjedésű bükk (Fagus sylvatica). A szikeseink egyik jellemző társulása pl. a cickóros puszta (as. Achilleo-Festucetum pseudovinae). Ebben karakter faj az Achillea asplenifolia és a Festuca pseudovina is. A két karakterfaj nevéből képezzük a társulás tudományos nevét is. Többféle társulás koncepció is létezik, mely a társulások határait és dinamikáját más-más módon értékeli.
1. Clements szerint: az élőhelyen hasonló környezeti háttér mellett ugyanazok a populációcsoportok élnek – „ mindig együtt fordulnak elő” - és ezáltal stabil fitocönozisokat építenek fel, mert egy niche grádiens mentén szerveződnek.
2. Gleason felfogása szerint: a populációk kapcsoltsága gyakran véletlenszerű, bármely időszakban az adott környezet formálja, ezért létrejöhet egy véletlenszerű „gyülekezet” is. A társuláshatárok gyakran elmosódnak, sokféle átmenet létezik.
A növénytársulások valóban dinamikus egységek, és bár határai sok esetben jól körvonalazódnak, idővel megváltoznak és a populáció közösségek kicserélődnek. A társulások dinamikájában egy térbeli és egy időbeli faktor is létezik. Térben a változás lehet horizontális, ilyen a mintázat és vertikális, mint a például a szintezettség. Az időbeni változás lehet periodikus és előremutató. Előbbire jó példa az erdők aszpektusainak váltakozása, utóbbira pedig a szukcesszió.
A szukcesszió
Ez a folyamat a növénytársulások egymásutániságát írja le. A növénytársulások váltása a környezeti tényezők megváltozása és a belső szerkezeti átrendeződések miatt következik be.
Jellemzői szerint a szukcesszió lehet:
- szekuláris szukcesszió, mely hosszú, földtörténeti léptékű, meg nem ismétlődő és a nagy klímatörténeti változásokat kíséri.
- biotikus szukcesszió, amely rövidebb idő alatt, változatlan makroklíma viszonyok mellett zajlik, és mindig egy ismétlődő folyamat. Együtt jár a környezet teljes átalakulásával, megváltozásával, melyeket
endogén okok (pl. humusz képződés),
exogén – allogén - okok (pl. folyóhordalék árvízek után)
váltják ki.
A szukcesszió kutatások atyja, F.I. Clements hat alapfolyamatot említ a szukcesszió során. Ezek a nudáció (terület lecsupaszodása), migráció (propagulumok bevándorlása), ökozis (a populációk idomulása az új környezethez), reakció (a környezetre való visszahatás eredményeképpen a populációk átalakítják a környezetüket), kompetíció (forrásokért versengés), stabilizáció (állandósulás) Minden biotikus szukcesszió sorozatnak van egy kezdeti szakasza, ez a pionír fázis és egy végső szakasza a klimax.
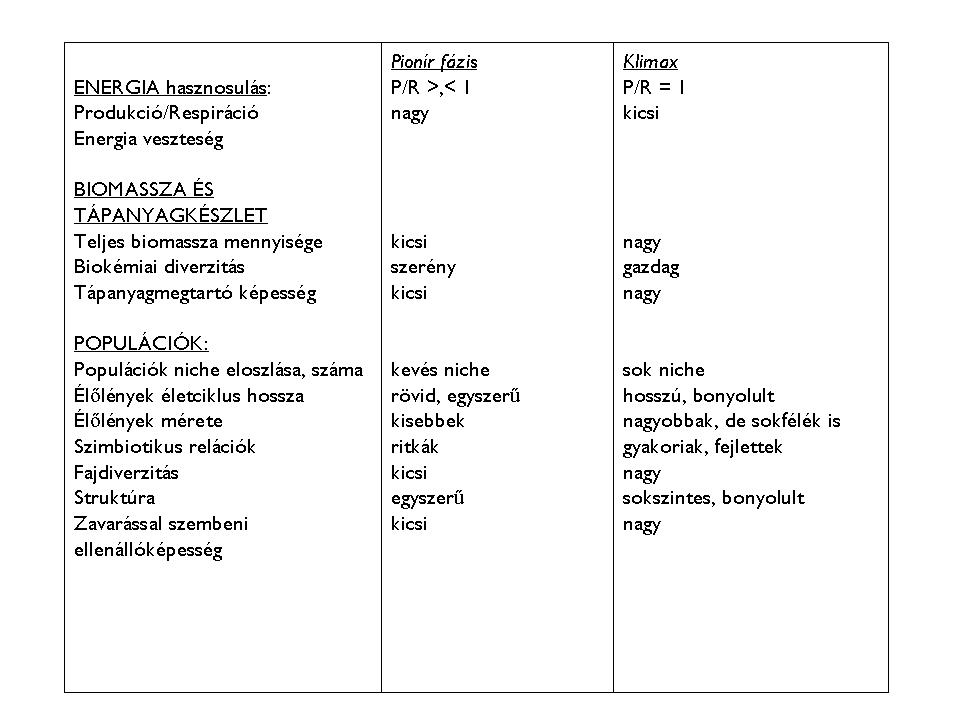
A két fázis között átmeneti szakaszok váltakoznak (szukcessziós szériesz), melyek egymástól az anyag- és energiagazdálkodásban egyaránt nagyon különböznek. A végső fázis a stabilitáshoz vezet, melyet egy dinamikus egyensúly szabályoz. A klimax stádium tehát a legnagyobb szervesanyag-termelést felmutató és legtöbb szintből álló növénytársulás mely az ökoszisztéma keretei között hatékony anyagforgalmat és gazdaságos energiafelhasználást valósít meg. A populációk közötti kapcsolatok igen sokrétűek, bonyolultak (1. Táblázat)
1. táblázat: A pionír és klimax társulások összehasonlító jellemzése (Odum, H.T. és Juhász Nagy Pál nyomán módosítva)
A szukcesszió formái
- primér (még nincs kialakult talaj): pl. morénák benépesülése a jégtakaró visszaszorulása után, zátonyok, kihűlt lávafelszínek (25. Ábra).
- szekundér (már van talaj, korábbi vegetáció maradványokból) pl. szántók, erdőirtások, magas árvízek utáni időszakok (26. Ábra).
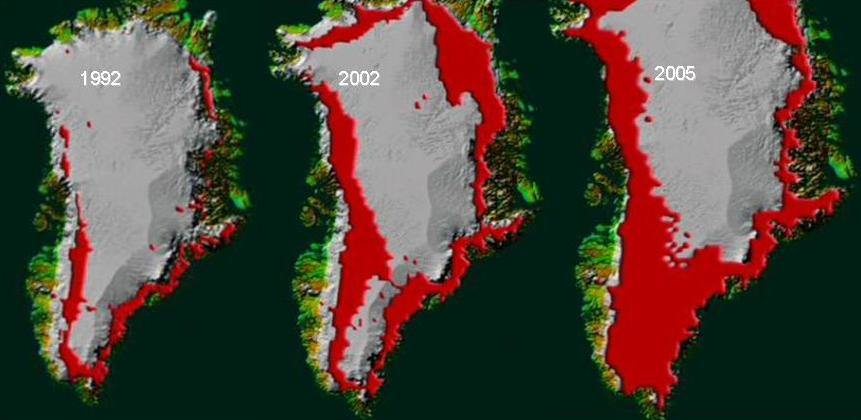
25. Ábra: Olvadó jégtakaró képe Grönlandon
Hazai viszonyok között az alföldi tájaink homok felszínein és a leomló löszfalakon figyelhető meg primér szukcesszió sor, melynek szemléletes leírása Fekete Gábor (1992) nevéhez fűződik. Azonos klímában tehát, de eltérő szubsztrátumon két markánsan eltérő vegetáció fejlődik ki: a klímazonális löszvegetáció és az extrazonálisnak tekinthető homoki vegetáció.
A homoki szukcesszió (lásd. a homoki növényzet a Duna-Tisza közén kisfilm)
- A homok durvaszemcsés szerkezetű üledék, mely rossz talajképző tulajdonságokkal rendelkezik.
- A szélsőséges talajviszonyok miatt rossz tápanyagellátás és vízgazdálkodás jellemző.
- Az itt élő fajok tágtűrésűek, többnyire nem specialisták.
- A populációk között a források felosztása egyenlőtlen.
- Homokon nincsen lineáris vegetációfejlődés: a nyílt gyep rendszerint hosszú életű.
- Nincs jellegzetes cserjés, a fajok sodródnak, a homok a „környékről” gyűjti be a többnyire tágtűrésű generalista fajokat.
- Nincs egyértelmű klimax társulás, vagy legalábbis nem tudjuk mi lehetett. Az a fás vegetáció (fehérnyáras-borókás), amely ma jellemzi a pannon homokhátakat semmiképpen sem hordozza a zárótársulás jegyeit..
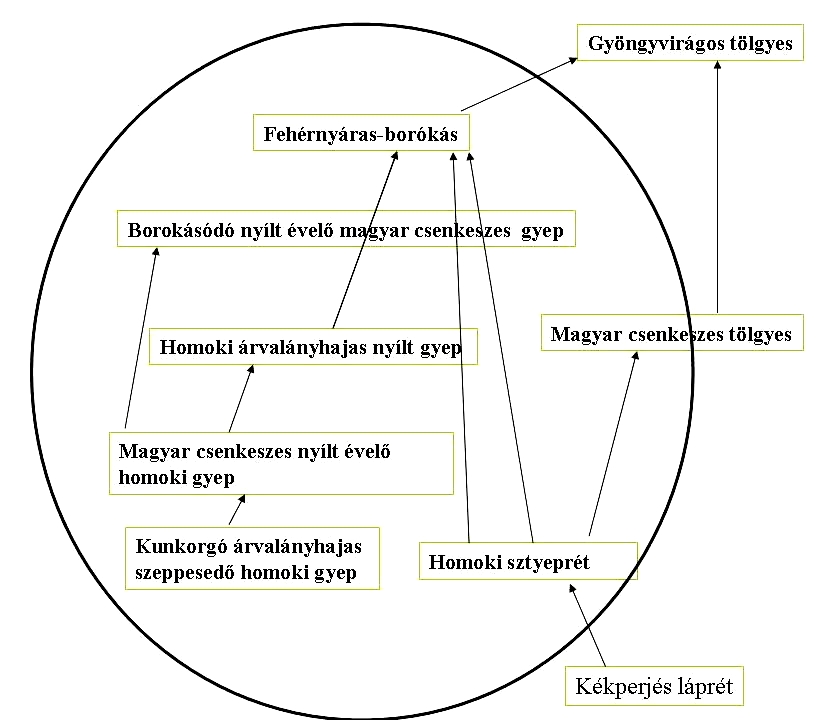
26. Ábra: A homokon zajló edafikus szukcesszió sémája (Fekete G. 1992. nyomán)
Homoki növényzet a Duna-Tisza közén (kisfilm)
A löszvegetáció fejlődése (lásd. a lösz növényzet a Duna mentén kisfilm)
- Jó talajképződési folyamatok miatt, a növényzet jó tápanyagellátás és vízháztartás mellett fejlődhet.
- A fajok kezdetben tágtűrésűek, de már a sztyepréttől kezdődően precíz niche felosztás valósul meg.
- Lineáris vegetációfejlődés jellemző, mely a pionír fázistól halad a klimax felé.
- Saját, löszcserjés, löszjelző cserjefajokkal.
- Egyértelmű, fajgazdag klimax társulás, mely nálunk a szubmediterrán jellegű tatárjuharos lösztölgyes.
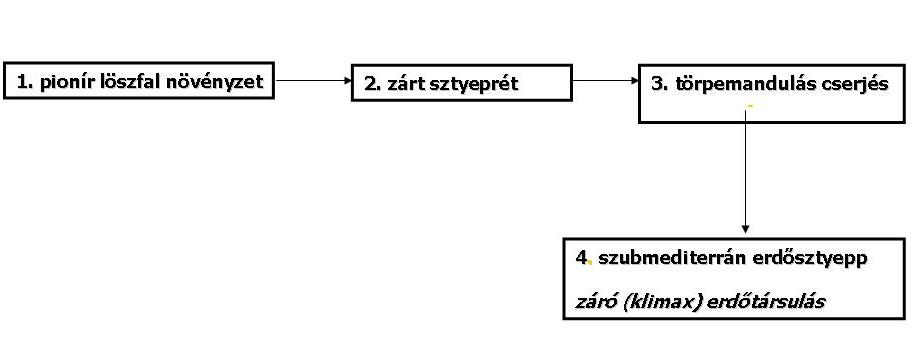
27. Ábra: A löszön zajló zonális szukcesszió sémája (Fekete G. 1992. nyomán)
Lösz növényzet a Duna mentén (kisfilm)
A löszterületeinket mára már többnyire művelés alá vonták, a felhagyott szántókon azonban a szekundér szukcesszió beindulásával újra kialakulhat a löszsztyep, ha a magkészlet még hordozza az életképes propagulumokat.

28. Ábra: Felhagyott szántó benépesülése egyéves fajokkal (Papaver rhoeas) egy alföldi löszháton.
Gyakran megfigyelhető, szekundér szukcesszió megy végbe a felhagyott bányák meddőhányóin is.

29. Ábra: Gánti (Vértes) bauxit bányászat után létrejött bányameddő felszíne ( forrás: http://www.karpat-medence.hu)
Irányát tekintve a szukcesszió lehet:
- progresszív vagy fejlődő, amely a klimaxállapot felé tart
- regresszív vagy hanyatló, mely leromló stádiumhoz, degradációhoz vezet, és végső soron a növénytársulások összeroppanását válthatja ki. Ilyenek a folyamatos erdőirtásnak kitett területek, ahol az erózió miatt lepusztult talajon az esetlegesen betetelepülő fajok hosszú ideig struktúra nélküli állományokat alakítanak ki. A mediterrán térségben többfelé figyelhető meg ez a jelenség.
A szukcesszió sebessége mindig függ:
- az aljzat tulajdonságaitól
- az éghajlati tényezőktől
- a propagulum forrásoktól való távolságtól
A térbeli szukcessziót nevezzük zonációnak is. A zonációban a szukcessziós fázisok gyakran egymás mellett és megjelennek. Ilyen például a folyóvízi- és az állóvízparti szukcesszió (28. 29. Ábra) .

30. Ábra: Folyóvízparti zonáció (Csűrös I. nyomán)
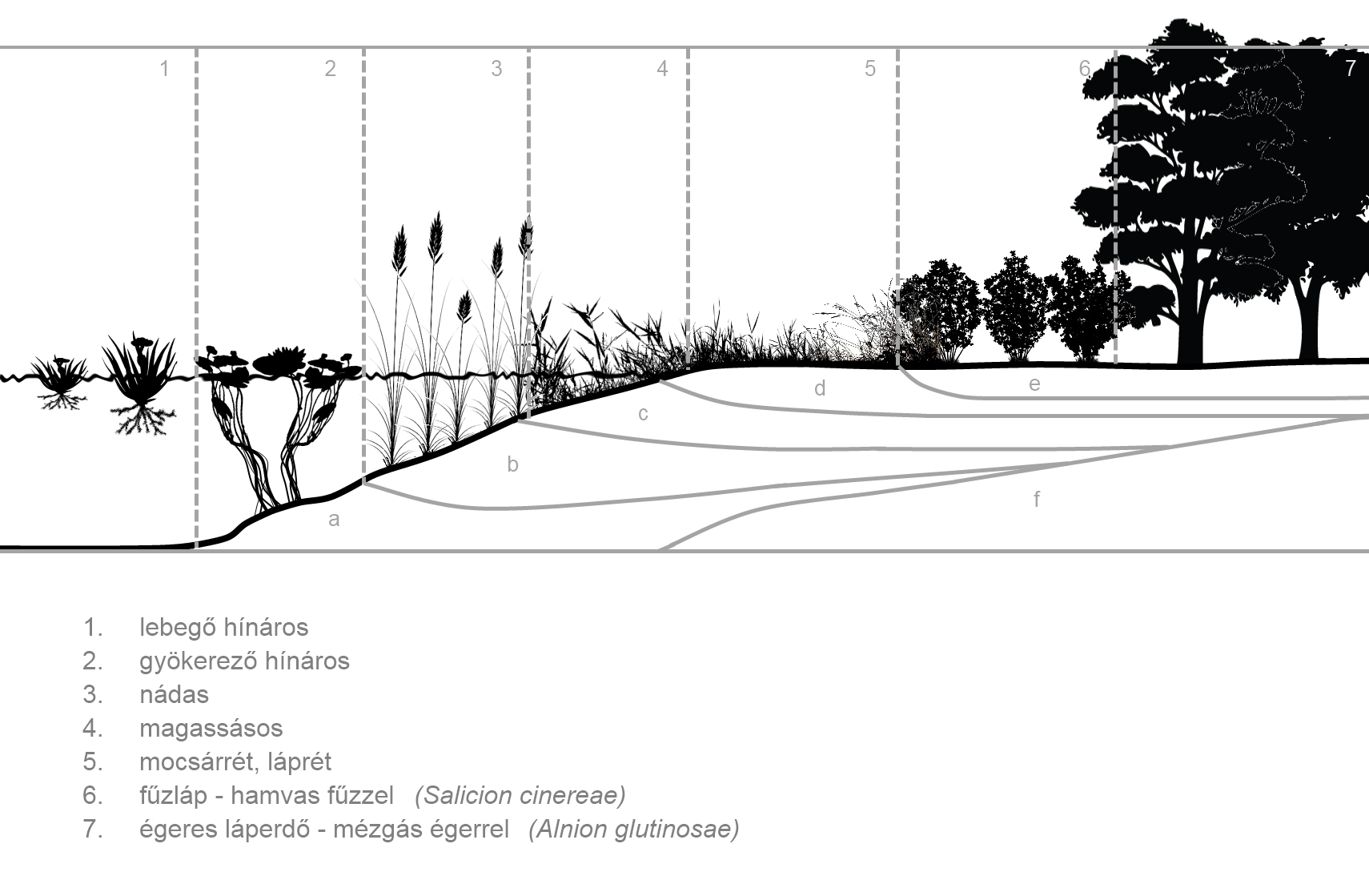
31. Ábra: Állóvízparti, tavi zonáció (Soó R. nyomán)
A szukcesszió nyomon követése bármely természetközeli vagy kultúr élőhelyen fontos lehet, ha a degradáció megelőzését kívánjuk elérni. Emberi tevékenységgel a progresszív vegetációfejlődési folyamatokat juttathatjuk előtérbe. A progresszív folyamatok révén az élőhelyek sokkal költségtakarékosabban tarthatók fönn és emellett egy élhető táj és környezet alakulhat ki.
Ellenőrző kérdések és feladatok
1. Definiálja a limitáló tényező fogalmát!
2. Mi a mikorrhiza? Sorolja föl a tanult mikorrhiza típusokat(4)!.
3. Írjon példafajt két allelopatikus hatású növényre.
4. A tanult populációk közötti kapcsolatokat írja be a kapcsolat típusától függően a megfelelő négyzetbe!
|
+/+ |
+/- |
|
-/- |
+/0 |
Válassza ki az alábbi kérdésekre a helyes választ úgy hogy akár több válasz is helyes lehet:
5. Mikor lép föl kompetíció két faj populációja között ?
a. Két magasabbrendű faj együttélésénél.
b. Bezárt niche-k esetén.
c. Diszjunkt niche-nél.
d. Niche átfedési zónában.
6. Mely fajok lehetnek jó pionírok?
a. A rovarmegporzásúak.
b. A széllel terjedő magvúak.
c. A szűktűrésű specialisták.
d. A vegetatívan is jól terjedők.
7. A szimbiotikus kapcsolatok jellemzői:
a. Pozitív interakció a partnerek között.
b. A klimax társulásokban rendszerint gyakoribb, mint a pionír társulásokban.
c. Gyakori alacsonyabb rendű és magasabb rendű fajok populélációi között.
d. Ideiglenes kapcsolat a populációk között.
8. Pontokba foglalva jellemezze és hasonlítsa össze a pionír és klimax növénytársulás fontosabb jellemzőit
|
|
pionír fázis | klimax |
|
Energiagazdálkodás |
||
|
Tápanyag ellátottság |
||
|
Biomassza tömeg |
||
|
Populáció struktúra |
9. Írja le az állóvízparti és folyóvízparti szukcesszió közötti alapvető különbségeket pontokba foglalva legalább négy szempont szerint.
| Szukcesszió jellemzői | állóvíz parton | folyóvíz parton |
Irodalomjegyzék
Cain, M., Bowman, V.D., Hacker, S.D. 2011. Ecology. (2nd ed). Sinauer Associates Inc.
Fekete G. (szerk.) 1985: A cönológiai szukcesszió kérdései. Biológiai Tanulmányok 12. Akadémiai Kiadó, Budapest.
Mátyás Cs. 1997: Erdészeti Ökológia. Mezőgazda kiadó, Budapest.
Fekete, G., Molnár, Zs., Horváth, F. 1997: A magyarországi élõhelyek leírása, határozója és a Nemzeti Élõhelyosztályozási Rendszer. Nemzeti Biodiverzitás-monitorozó Rendszer II. MTA-ÖBKI - KTM kiadás, Budapest.
Pásztor E., Oborny B. (szerk.) 2007: Ökológia. Nemzeti Tankönyvkiadó, Budapest.
Simon T. (szerk.) (2001): Növényföldrajz, Társulástan és Ökológia. Tankönyvkiadó, Budapest.
Török p., Tóthmérész B. (2006): Növényökológiai alapismeretek. Kossuth Egyetemi kiadó, Debrecen.
Turcsányi G. (szerk.) 1998: Mezőgazdasági növénytan. Mezőgazdasági Szaktudás Kiadó, Budapest. 2. kiadás
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028