Szövettan (Hisztológia)
Szerző: Erős-Honti Zsolt
A növényi szövetekről általában
A szövet fogalma az azonos eredetű, hasonló felépítésének köszönhetően hasonló működésre specializálódott sejtek csoportját jelöli. A növényi szervezet jellegzetes fejlődése – a differenciálódó sejtek meghatározott elköteleződési rendszere – miatt az egyes szövetek kialakulása meghatározott sejtcsoportokra, sok esetben konkrét sejtekre vezethető vissza. A differenciálódott sejtek ezért – ontogenetikus (egyedfejlődési) történetük alapján – hierarchikus rendszerbe csoportosíthatók. Az egy konkrét feladatot ellátó sejtek csoportjait (pl. a sztómakomplex sejtjeit) egyszerű szöveteknek nevezzük, melynek tagjai a legtöbb esetben a megfelelő szövetet kialakító osztódó sejtcsoport (osztódószövet) egyetlen sejtjéből jönnek létre. Az egyszerű szövetek szövetekké (pl. epidermisz, xilem) állnak össze. A hasonló funkciót ellátó (következésképpen hasonló elhelyezkedésű és szerkezetű) szövetek összességét nevezzük szövetrendszernek (pl. a bőrszövetrendszer kategóriájába tartozik az epidermisz, a rizodermisz és a periderma – részletesen lásd később!).
A szöveti szintű specializáció a növények evolúciója során a szárazföldre lépéssel vált szükségessé. A tengerekből kilépő növényvilág tagjai közül az új élettérben csak azok az egyedek maradhattak életben, amelyek sejtjei között a munkamegosztás (és az ezzel párhuzamosan fellépő alaki változatosság) már elérte a szövetek szintjét. (A mohák csoportjainak körében a szárazföld meghódítása szintén a sejtek differenciálódását vonta magával, azonban ez még nem haladta meg a telepes testszerveződésre jellemző álszövetek szintjét. Ezzel magyarázható, hogy az ősi mohacsoportokból nem alakultak ki összetettebb testfelépítésű élőlények: az evolúció oldalágainak tekinthetők.)
Valódi szövetekkel, szövetrendszerekkel a harasztok, a nyitvatermők és a zárvatermők (tehát a hajtásos vagy edényes növények) rendelkeznek.
A növényi szövetek csoportosítása és kapcsolatrendszere
A növényi szöveteket két nagy csoportba sorolhatjuk: az osztódószövetek (merisztémák) és az állandósult szövetek kategóriájába. Az osztódószövetek folyamatos mitotikus gyarapodásuk révén biztosítják az egyed sejtszámának növekedését. Az osztódószövet sejtjeinek két-két leszármazottja közül az egyik differenciáltsági szintje a kiindulási sejtével azonos marad, míg a másik a további differenciálódás útjára lép. A differenciálódás jelentheti azt, hogy egy már elkötelezettebb, de még mindig osztódni képes merisztémasejt lesz belőle vagy elveszítve mitotikus aktivitását állandósult szöveti sejtté alakul (tehát kilép a sejtciklusból az ún. G0-stádiumba). Egyes esetekben előfordulhat, hogy a növényi szervezet fejlődésével egyes állandósult szöveti sejtek visszanyerik osztódóképességüket (dedifferenciálódnak, visszalépnek a sejtciklusba), ekkor azonban már nem tekinthetjük őket állandósult szöveteknek: ismét merisztémává alakulnak át.
A legkevésbé elkötelezett sejtekből álló osztódószövetek a promerisztémák (ősmerisztémák vagy iniciálisok) (1. ábra). Az ősmerisztéma-sejtek leszármazottai még mindig osztódóképes sejtek: ezek az elsődleges merisztémák (primer merisztémák). Ezeknek a sejteknek a mitotikus aktivitása már differenciálódott, állandósult szöveti sejteket hoz létre, az elsődleges szöveteket. Abban az esetben, ha a növény fejlődése azt megköveteli (azaz genetikai programjában kódolt), az elsődleges szöveti elemek visszanyerhetik osztódóképességüket (pl. a szár vagy a gyökér másodlagos vastagodása esetén – lásd később!). Az így kialakuló osztódószövet neve másodlagos (szekunder) merisztéma (pl. a parakambium). A másodlagos merisztémák aktivitása is állandósult szövetekkel gyarapítja a növényi testet; ezek a másodlagos szövetek (pl. a periderma).
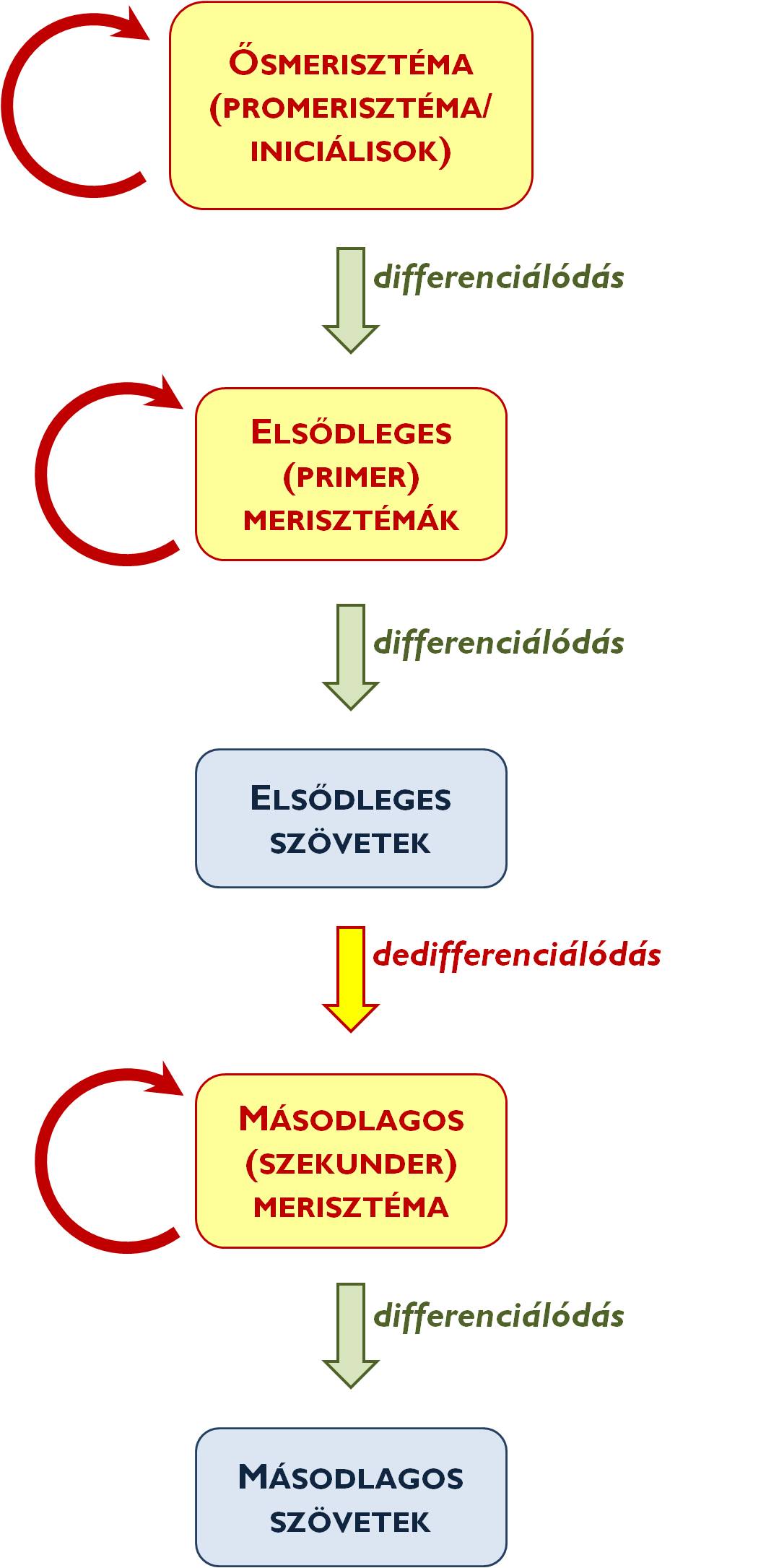
1. ábra. A növényi szövetek leszármazási kapcsolata.
Az állandósult szövetek három nagy szövetrendszerre tagolhatók. A növényi testet legnagyobb részben az alapszövetrendszer építi fel, amelynek tagjai igen változatos feladatokra (fotoszintézis, raktározás, kiválasztás) módosulhatnak. A bőrszövetrendszer feladata a növényi szervezet elhatárolása, védelme, ugyanakkor a kapcsolat biztosítása a szervek belső állománya és a külvilág között. A szállítószövetrendszer a növényi tápanyagokat (víz és ásványi anyagok) valamint a szerves vegyületeket (cukrok, aminosavak, egyes hormonok) juttatja el a szervezet egyik részéről a másikba.
Az osztódószövetek (merisztémák)
Szemben az állati szervezettel, amelynek legtöbb sejtje osztódásra képes, a növényi test gyarapodása meghatározott területekre, a merisztémákra korlátozódik. Az osztódószöveti sejtek mitózisa hozza létre az összes többi sejtet, amelyek később a differenciálódás útját bejárva működő, állandósult szövetekké alakulnak. A merisztéma-sejtek citológiai jellemzői egyértelműen erre az egyetlen funkcióra utalnak. A viszonylag kisméretű, többnyire izodiametrikus (tehát a tér minden irányában többé-kevésbé azonos méretű, de a sejtek egymáshoz nyomódása miatt sokszöglapokkal határolt) sejtek sejtmagjának térfogata igen nagy a sejtplazma mennyiségéhez (illetve a teljes sejttérfogathoz) képest. A sejtek még differenciálatlanok: sejtfaluk vékony, tágulékony elsődleges sejtfal, plazmájukban több, kisméretű vakuolum helyezkedik el, plasztiszaik – amennyiben van bennük színtest – proplasztiszok.
A merisztémákat többféleképpen is csoportosíthatjuk.
I. Eredet és elkötelezettség alapján:
- Promerisztéma: a legkevésbé elkötelezett, iniciális sejtekből álló osztódószövet.
- Elsődleges merisztéma: a promerisztéma differenciálódásával kialakuló osztódószövet. Működése az elsődleges szöveteket hozza létre.
- Másodlagos merisztéma: elsődleges (állandósult) szöveti sejtek dedifferenciálódásával kialakuló osztódószövet. Működése másodlagos szöveteket hoz létre.
II. Elhelyezkedés alapján:
- Csúcsmerisztémák: a növény hossztengely mentén megfigyelhető növekedését biztosító, a növényi test pólusain elhelyezkedő osztódószövetek: a hajtáscsúcs és a gyökércsúcs. (E két merisztématípus folyamatos aktivitásának köszönhetik a „növények” magyar megnevezésüket is: az egyed élete végéig tartó osztódásuk miatt hívjuk őket „növekvő lények”-nek.)
- Interkaláris merisztémák: a szervek másodlagos megnyúlását szolgáló osztódószövetek, amelyek nem csúcsi helyzetben, hanem állandósult szövetek közé ékelődve fordulnak elő a növényben. Pl. a szalmaszár vagy a tőszár internódiumaiban, a pázsitfűfélék levelének alapi régiójában.
- Oldalmerisztémák: a szervek vastagságbeli gyarapodását okozó, a szerv keresztmetszetén perifériális helyzetben elhelyezkedő osztódó szövetek. Ebbe a csoportba tartozik a szállító- és alapszöveteket létrehozó vaszkuláris ill. hullámos kambium, valamint a másodlagos bőrszövetet (a peridermát) kialakító parakambium (fellogén).
Az egyes szerveket kialakító elsődleges és másodlagos merisztémák típusait és elhelyezkedését a megfelelő szervszövettani fejezetek ontogenezissel foglalkozó részeinél tárgyaljuk részletesen.
A bőrszövetrendszer
A növények szervezetét a külvilág felé a bőrszövetrendszer elemei határolják: ezek a szövetek felelősek a növényi test integritásának kialakításáért. A testfelszíni elhelyezkedésnek megfelelően e szövetrendszer feladata az elhatárolás mellett a külvilág és a szervezet belseje közti kapcsolat fenntartása is. Alapvető funkciói ennek megfelelően: a szervezet védelme a mechanikai, kémiai és biológiai (patogén) hatásokkal szemben, az anyagfelvétel, a párologtatás és a gázcsere biztosítása. A bőrszövetrendszer emellett raktározó és kiválasztó funkciót is elláthat.
A bőrszövetrendszert – állandósult szövet révén – további osztódásra nem képes, vastag másodlagos sejtfallal és nagyméretű vakuolummal rendelkező, a sejt térfogatához képest kisméretű sejtmagot tartalmazó sejtek építik fel. A védelmi funkció ellátása érdekében a sejtek között szorosak a kapcsolatok (tehát nincsenek sejtközötti járatok), gyakran ezt tovább erősíti az is, hogy a szomszédos sejtek igen nagy felületen, hullámos lefutású sejtfallal illeszkednek egymáshoz (a sejtek a puzzle elemeihez hasonló alakúak). A bőrszövet sejtjei közti kapcsolatok erősebbek, mint az alattuk levő alapszöveti sejtekkel létesített kapcsolataik, ezért lehet a bőrszövet sejtjeit – többé-kevésbé – könnyen lehúzni a szervek felszínéről (nyúzatkészítés). A sejtek nagy felületűek, laposak, többnyire egy sejtréteget alkotnak. Bennük zöld színtesteket csak kivételes esetben találunk (pl. víz alá merült, vékony levelek bőrszövete esetében, illetve a sztómák zárósejtjeiben). (Megjegyzendő azonban, hogy raktározó leukoplasztiszok előfordulhatnak bennük.) A sejtek külvilág felé eső sejtfala gyakran erősen megvastagszik: kutin-, szuberin- és/vagy viaszréteg lerakódása miatt.
A gyökér elsődleges bőrszövete: a rizodermisz
A gyökér egyik alapfeladata a növényi tápanyagok (víz és ásványi anyagok) felvétele a talajból. Ezt a szerepet – a zárvatermőknél – a gyökér elsődleges bőrszövetének (rizodermisz) speciális bőrszöveti képletei, a gyökérszőrök látják el (+info). A gyökérszőrök a hajszálgyökerek felszívási zónájában (lásd később!) fordulnak elő. Nem valódi növényi szőrök, mindössze a bőrszöveti sejtek kesztyűujj-szerű nyúlványai, és nem különülnek el sejtfallal az őket kialakító bőrszöveti sejttől. Kialakulásuknak köszönhetően jelentősen megnövekszik a bőrszövet anyagfelszívó felülete.
A rizodermiszt az alább tárgyalandó epidermisztől a gyökérszőrök jelenléte mellett az is megkülönbözteti, hogy általában nem találunk benne sztómákat, (valódi) növényi szőröket és kutikularéteg is csak ritkán fedi.
A valódi léggyökerek bőrszövete: a velamen radicum
A hajtás elsődleges bőrszövete: az epidermisz
A korábban megismert bőrszöveti funkciók nagy része a hajtást borító elsődleges bőrszövethez, az epidermiszhez köthető. A hajtáson (azaz a leveleken és a fotoszintetizáló szárakon) található gázcserenyílások (sztómák) biztosítják a párologtatást és a gázcserét. A sejteket gyakran vastag kutikularéteg fedi, ami a védelmi funkciót szolgálja, hasonlóan az epidermisz szőrképleteihez. A különféle mirigyek, mirigyszőrök a kiválasztást teszik lehetővé.
A gázcserenyílások (sztómák)
A növényi test belső teréből a vízgőz, a légzés során keletkezett CO2 illetve a fotoszintézis termelte O2 a bőrszövet szabályozottan nyitható és zárható nyílásain, a gázcserenyílásokon (sztóma) át távozik. Itt veszi fel a növény a fotoszintézishez szükséges CO2-ot és a légzéshez szükséges O2-t is. A gázcserenyílások működését többféle, speciális felépítésű sejt teszi lehetővé, ezért sztómák helyett helyesebb sztómakomplexekről beszélni.
A sztómakomplex felépítésében minden esetben részt vesz két zárósejt, amelyek a sztóma szabályozható nyílását, a légrést veszik körül. A légrés a bőrszövet alatti alapszövet egy tág sejtközötti üregébe, a légudvarba vezet. A zárósejtek alakja a fajok többsége esetében babszemre emlékeztet, de a pázsitfűfélék (Poaceae) családjában súlyzóhoz hasonló megjelenésűek. A zárósejtek jellegzetessége, hogy bennük kloroplasztiszok fordulnak elő (tehát fotoszintézisre képesek). Sejtfaluk egyenlőtlen vastagodást mutat: a légrés melletti szakaszokon vastagabb, mint a zárósejt egyéb felületein. A zárósejtek sejtfalára emellett kutikularéteg is rakódik. Ennek vastagsága a légrés felőli oldalon erőteljesebb, ráadásul a légréssel érintkező felületen egy kutikula-kiemelkedés (kutinléc) segíti a légrés tökéletes záródását. (Egyes fajok esetében a zárósejtnek mind a külvilág felé, mind a belső, szövetek fel eső oldalán találunk kutinlécet.)
A zárósejtek működése
A zárósejt működésének alapja a sejten belüli nyomás (turgor) változása. A légrés kinyílását megelőzően a zárósejtek – aktív transzporttal – K+-ionokat vesznek fel a környezetükből, ami a sejten belüli ionkoncentráció megemelkedéséhez vezet. Ennek köszönhetően megnő a sejt vízfelvevő képessége és ozmózissal víz áramlik a sejtek belsejébe. A megnövekvő vízmennyiség hatására fokozódik a sejtplazma sejtfalra gyakorolt (turgor)nyomása, ami – a léggömbbe fújt levegőhöz hasonlóan – feszíti sejtfalat: a sejtfal tágulni kezd. Az egyenlőtlen sejtfalvastagodásnak köszönhetően azonban ez a tágulás a sejt különböző felületein különböző mértékű lesz: a légrés két oldalán minimális, ettől átellenesen pedig maximális. Az egyenlőtlen tágulás a zárósejtek alakváltozását (meghajlását) okozza: ezzel kinyílik a légrés.
A záródás folyamata ezzel ellentétes mechanizmussal valósul meg: a sejt leadja a környezetébe a K+-ionokat, a lecsökkenő belső koncentráció vízvesztéshez, következésképpen turgorcsökkenéshez vezet. A zsugorodó zárósejtek alakváltozásának köszönhetően zárul a légrés.
Animáció a sztómák működéséről: az animáción a sztómazárósejtek turgornövekedés okozta alakváltozása látható.
A zárósejtek működésének szabályozása
A legtöbb nemzetség zárósejtjeit csak az epidermisz általános bőrszöveti sejtjei övezik; ilyenkor anomocitikus sztómáról beszélünk (pl. Cyclamen /ciklámen/, Ranunculus /boglárka/, Cornus /som/ vagy Iris /nőszirom/ nemzetségekben) (2. ábra). Máskor a zárósejtek működését a többi bőrszöveti sejttől eltérő megjelenésű melléksejtek segítik. Amennyiben két melléksejt helyezkedik el a zárósejtekkel párhuzamosan a sztómakomplexben, a sztóma paracitikus (pl. Rubiaceae /galajfélék/, Magnoliaceae /liliomfafélék/). Ha a melléksejt-pár síkja merőleges a zárósejtekre, diacitikus a sztóma (pl. Lamiaceae /ajakosok/ vagy Caryophyllaceae /szegfűfélék/ családja). Az anizocitikus sztómakomplex 3-5, pl. két kisebb és egy nagyobb méretű melléksejtet tartalmaz (pl. a Begonia /begónia/, Sedum /varjúháj/ fajok vagy a Brassicaceae /keresztesvirágúak/ családja esetében). Ha a zárósejt körül több, sugárirányban álló melléksejt van, a sztóma aktinocitikus (pl. Anacardiaceae [szömörcefélék]). Gyűrű alakban rendeződnek a melléksejtek a ciklocitikus sztómakomplex esetében (pl. egyes Austrobaileya és Baccharis /borfa/ fajok). A felsorolt alapvető típusok mellett számos további melléksejt-elrendeződés is elképzelhető (pl. a hat melléksejtes hexacitikus vagy a több, különböző körkörösen rendeződő melléksejtet tartalmazó heliocitikus sztómakomplex.)
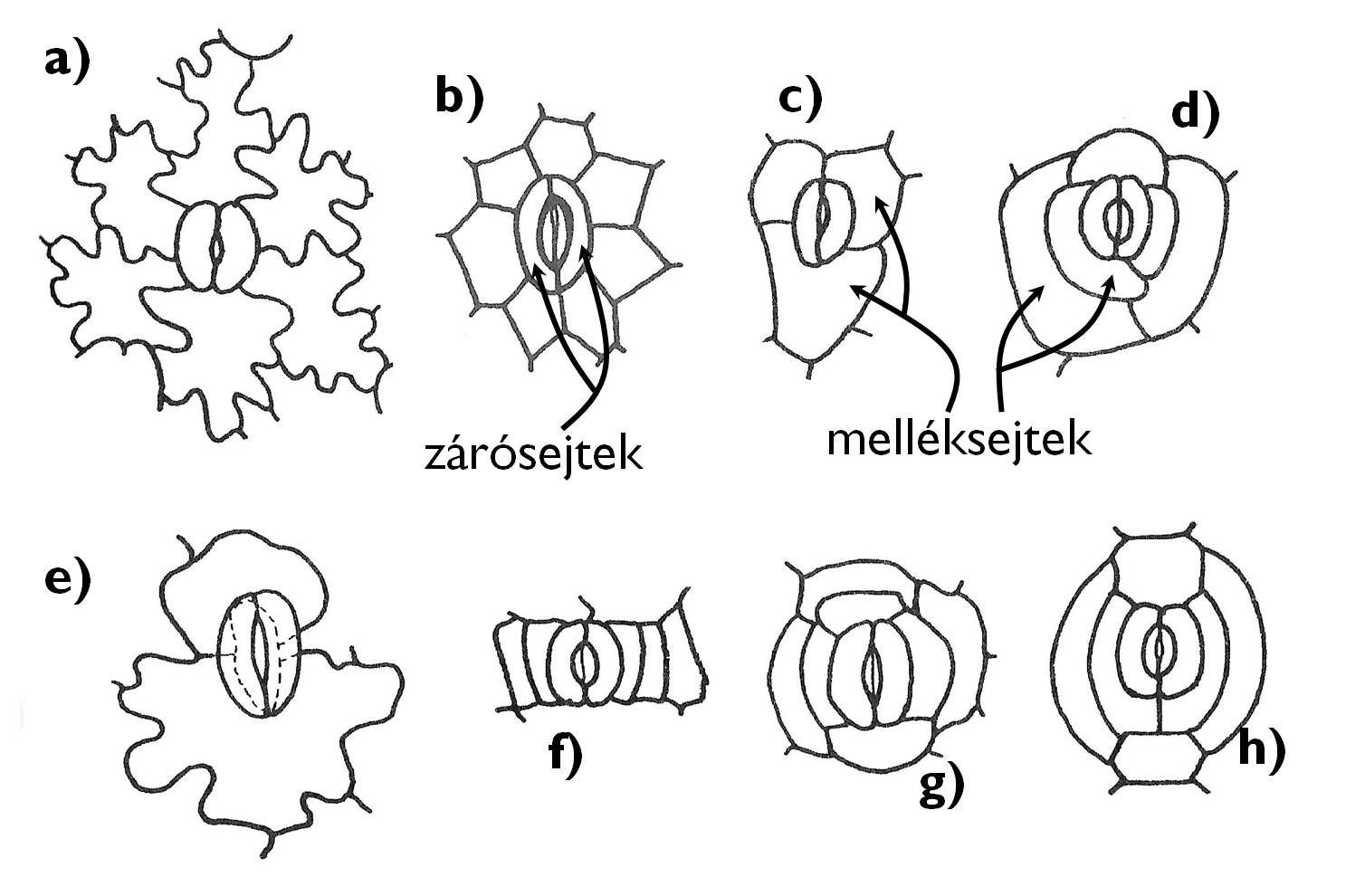
2. ábra. A gyakoribb sztómatípusok.
a) anomocitikus; b) aktinocitikus; c) anizocitikus; d) heliocitikus; e) diacitikus; f) paracitikus; g) ciklocitikus; h) hexacitikus.
(Gyurján nyomán)
A kutikula
A hajtás bőrszövetének egy tipikus sajátossága az epidermiszsejtek felszínén kialakuló kutikularéteg. A kutikula két legfontosabb vegyületcsoportja a kutikula alapállományát (mátrixát) képező, a vízben oldhatatlan kutin és a különböző viaszok. A viaszok különálló réteget is alkothatnak a kutikula felületén (epikutikuláris viaszok), de egy jelentős részük a mátrixba ágyazódik bele (intrakutikuláris vagy kutikuláris viaszok).
A kutikula rétegzett szerkezetű. A legkülső réteget az epikutikuláris viaszok alkotják (viaszréteg), ami hamvassá, bársonyos fényűvé teszi a szerv felszínét. Elsődleges szerepe a kutikulán keresztül történő vízleadás (kutikuláris párologtatás) mérséklése. A viaszréteg anyagai a kutikula repedésein át jutnak a felületre, ahol vagy összefüggő, amorf réteget képeznek, vagy többé-kevésbé kristályos formában válnak ki fonalakat, rövid pálcikákat, lemezeket alkotva.
A kutikula elsődleges szerepe a védelem. A vastag kutikulájú hajtásrészek merevek, kemények, ellenállóbbak a mechanikai hatásokkal, sőt akár a növényevő rovarok rágásaival szemben is. Vízlepergető hatása révén az átázástól is védi a növényi szervet, ugyanakkor gátolja a vízvesztést is. A kutikula egyes kémiai hatásokkal szemben is védelmet nyújthat.
A kutikula egyes biogén hatásoktól is védi a növényt. Kórokozók (gombák, baktériumok) támadása esetén, amikor a patogén bontóenzimei segítségével próbál áthatolni a hajtást borító bőrszöveten, enzimei elsőként a kutikularéteget kezdik el lebontani. Az elemésztett kutikulából olyan anyagok szabadulnak fel, amelyek gátolhatják a kórokozó szervezet további növekedését. A lebomló kutikulából felszabaduló más vegyületek jelmolekulákként (hormonhatású anyagokként) a növény védelmi rendszerét is aktiválhatják (pl. a felszabaduló oligoszacharidok).
Az epidermisz függelékei
A gyökér elsődleges bőrszövetével szemben az epidermisz valódi, az epidermiszsejtektől sejtfallal elkülönülő valódi növényi szőröket (trichómákat) hordozhat (3. ábra). Ezek szerepe és felépítése igen változatos lehet.
A legáltalánosabb esetben egyszerű kiemelkedéseket találunk az epidermiszsejtek felületén. Ez az ún. papillás epidermisz valójában még nem tekinthető szőrözöttnek. A felületnövelő képleteken megtörő fény bársonyos, selymes fényűvé teszi a szerveket – leggyakrabban a takaróleveleken fordul elő (pl. a Viola /árvácska/ vagy a Saintpaulia /afrikai ibolya/ pártáján).
Az epidermiszt többnyire egy- vagy többsejtű fedőszőrök borítják. Az utóbbiak egy ún. talpsejttel ágyazódnak a többi bőrszöveti sejt közé, és ehhez a talpsejthez kapcsolódnak a szőrképletet alkotó egyéb sejtek. A csillag- ill. pikkelyszőrök esetében (pl. az Elaeagnus angustifolia /keskenylevelű ezüstfa/ epidermiszén) a több talpsejtből álló nyélrészhez számos sugársejt kapcsolódik lapos, lemezszerű megjelenésű szőrt hozva létre. (A csillagszőrök esetében a sugársejtek száma jóval alacsonyabb, hosszuknak legalább feléig szabadon állnak, míg a pikkelyszőrök sugársejtjei szinte teljes hosszukban egymáshoz nőnek). Gyakori, hogy a fedőszörök tengelye elágazik, emeletes szőrképletet alkotva (pl. Lavandula angustifolia /levendula/, Verbascum phlomoides /szöszös ökörfarkkóró/). A fedőszőrök sejtjei sok esetben kialakulásukat követően elpusztulnak, és a vastag sejtfalon belüli üreget levegő tölti ki; ebben az esetben a szőrökkel borított szerv felszíne ezüstösen csillogó, fehéres színű.
A fedőszőrök feladata igen sokrétű. Mechanikai akadályt képezve csökkenthetik a növényevő rovarok kártételét. A szőrök közé szoruló levegő miatt hőszigetelő réteget hozhatnak létre a hajtásfelszínen – pl. a magashegységek növényei esetében (pl. Leontopodium alpinum /havasi gyopár/). Ugyanakkor az elhalt, világos szőrök visszaverik a levélfelületet érő fény egy részét (azaz növelik az albedóját). Ez meleg, napos élőhelyeken lehet előnyös, mivel megvédi a növényt a túlmelegedéstől (pl. a mediterrán Cerastium tomentosum /molyhos madárhúr/ esetében). Hasonlóan nyújt védelmet az erős szőrözöttség olyan élőhelyeken (pl. magashegységekben) is, ahol nagyobb a közvetlen (direkt) fény aránya a talajfelszínre jutó sugárzásban, ezáltal nagyobb a káros UV-komponens terhelő hatása is.
A szőrök között megrekedő levegő hamar telítődik vízgőzzel, ami csökkenti a hajtásfelület párologtatását. Ezért a fedőszőrök előnyt jelenthetnek a száraz, kevés felvehető vizet tartalmazó talajon élő növények esetében is. Ugyanakkor egyes broméliafélék (Bromeliaceae család) levélrozettát alkotó leveleit vízfelszívó pikkelyszőrök fedik, amelyek segítségével ezek a fán lakó fajok a levélrózsa ciszternájában felgyűlő csapadékvízből, vagy akár a vízgőzzel telt levegőből illetve a lehulló esőcseppekből képesek fedezni folyadékigényüket.
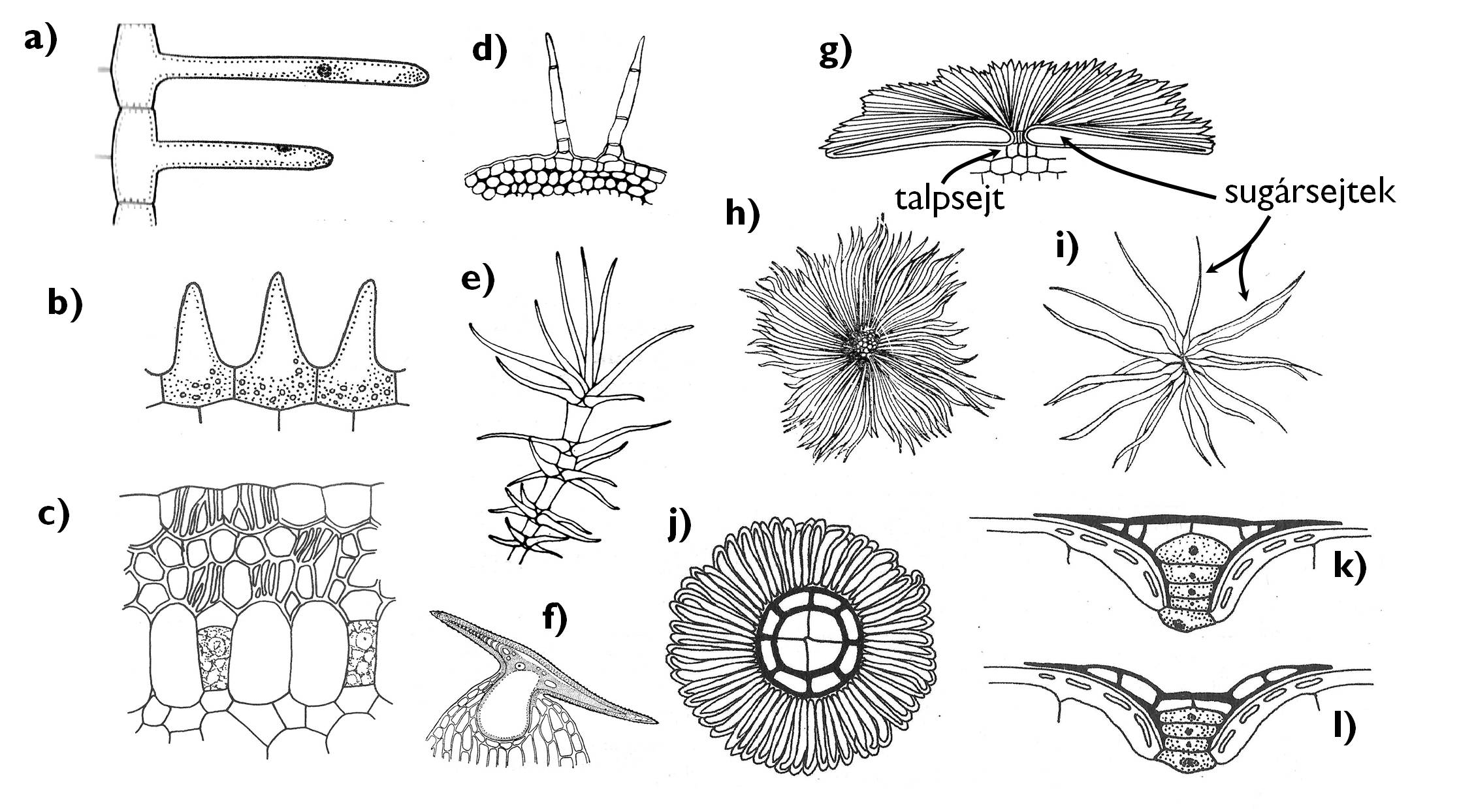
3. árba. A bőrszöveti függelékek.
a) gyökérszőr; b) papillák; c) velamen radikum; d) többsejtű, egyszerű fedőszőrök; e) emeletes fedőszőr (Verbascum); f) kapaszkodószőr ( Humulus); g) pikkelyszőr oldalnézetben; h) pikkelyszőr felülnézetben (Elaeagnus); i) csillagszőr (Elaeagnus); j-l) vízfelszívó pikkelyszőr felülnézetben (j), vízzel telt (k) és kiürült (l) állapotban (Bromeliaceae).
(a-b: Jacob és mts. nyomán, c: Gyurján nyomán, d-f: Kárpáti nyomán, g: Jacob és mts. nyomán, h-i: Sárkány és Szalai nyomán, j-l: Jacob és mts. nyomán)
A fedőszőrök szerkezetének, funkciójának megváltozásával egyéb szőrtípusok is kialakultak a növényvilágban az evolúció folyamán. Vastag sejtfallal rendelkező, merev fedőszőr a serteszőr (pl. a Boraginaceae /érdeslevelűek/ vagy a Cucurbitaceae /tökfélék/ család tagjain általános), feladata a növények fokozott védelme a rovarrágással szemben. Horgas, belülről zárvánnyal is merevített fedőszőröket találunk a komló (Humulus lupulus) hajtástengelyén: ezeknek a kapaszkodószőröknek a segítségével rögzül a szárkacs a támasztékán (a benne található zárvány miatt cisztolitszőrnek is nevezik). A magvak felszínét borító epidermisz (maghéj) is hordozhat szőröket (repítőszőrök), amelyek a magok széllel történő terjedését (anemochoria) segítik elő (pl. a Salix /fűz/, Populus /nyár/, Gossypium /gyapot/ nemzetségek magjain).
Az epidermisz kiválasztó funkciója: a növények felszíni kiválasztó képletei
A kiválasztás (szekréció) kifejezés azt a komplex folyamatot jelöli, melynek során a sejt eltávolít egyes anyagokat élő részeiből (protoplasztjából), vagy annak valamely részén elkülöníti azokat. A növényi szervezetnek lényegében valamennyi sejtje képes olyan anyagok előállítására, amelyek a sejten belül, az extracelluláris térben vagy akár a szervezeten kívül felhalmozódva látják el feladatukat. Ennek ellenére a szekréció jelenségét elsősorban a speciális, kimondottan kiválasztásra szolgáló szöveti elemekhez szokás kötni. Ilyen kiválasztó funkciót az epidermisz és a kiválasztó alapszövet végezhet. Ennek alapján felszíni és belső (endogén) kiválasztó struktúrákat különböztethetünk meg. Az előbbieket növényi mirigyeknek is nevezzük.
A mirigyeket alkotó sejtek vagy maguk állítják elő a leadott vegyületeket (mirigyszőrök, ozmofórák), vagy csak egyszerűen rajtuk keresztül jutnak a külvilágba a kiválasztott anyagok (pl. hidatódák, nektáriumok, sómirigyek). A leadott anyag lehet hidrofil (pl. nyálkaanyagok, nektár, sók) vagy hidrofób (pl. illóolajak) természetű. A kiválasztósejtekre minden esetben jellemző, hogy protoplazmájuk sűrű, a kiválasztott anyag természetétől függően egyes sejtalkotók mennyisége felszaporodik a sejtjeikben (pl. a nyálkaanyagot termelő sejtekben a Golgi-készülék, a lipideket előállító sejtekben a SER).
Az illatanyagok előállítása: az ozmofórák
Számos valódi kétszikű (Rosopsida) családban előfordulnak epidermális kiválasztó függelékek, ún. mirigyszőrök (4. ábra). E változatos felépítésű anatómiai képletek közös jellemzője, hogy kiválasztósejtjeik váladéka a külvilág felé néző kutikula rétege alatt halmozódik fel és a kutikula sérülése révén kerül a külvilágba. A kiválasztósejtek epidermiszre merőleges falaiba övszerűen szuberin vagy kutin rakódik le, a sejtfalban folyó (apoplasztikus – l. később!) transzport megakadályozása céljából. A kiválasztósejtek minden esetben elkülönülnek az epidermisz többi sejtjétől. A mirigyek szerkezete igen változatos: lehetnek egy- vagy többsejtűek, nyelesek vagy ülők (a Pelargonium [muskátli] nemzetség esetében mindegyik előfordul). Az ajakosok családjában (Lamiaceae) az epidermisz síkjába behúzódó ún. mirigypikkelyeket találunk, amelyek egy nagyobb nyaki sejtből, és az ezen ülő nyolc, illóolajat előállító kiválasztósejtből állnak. A mirigyszőrök illóolajak mellett egyéb hidrofób anyagokat is előállíthatnak: más terpenoidokat (gyantát, balzsamokat) vagy flavonoidokat. A leadott vegyületek számos célt szolgálhatnak: távol tarthatják a növényevőket, csalogathatják a megporzókat vagy ragadós természetük révén akár egyes termések terjedését is segíthetik.
Egy védelmi funkciójú mirigysejt: a csalánszőr
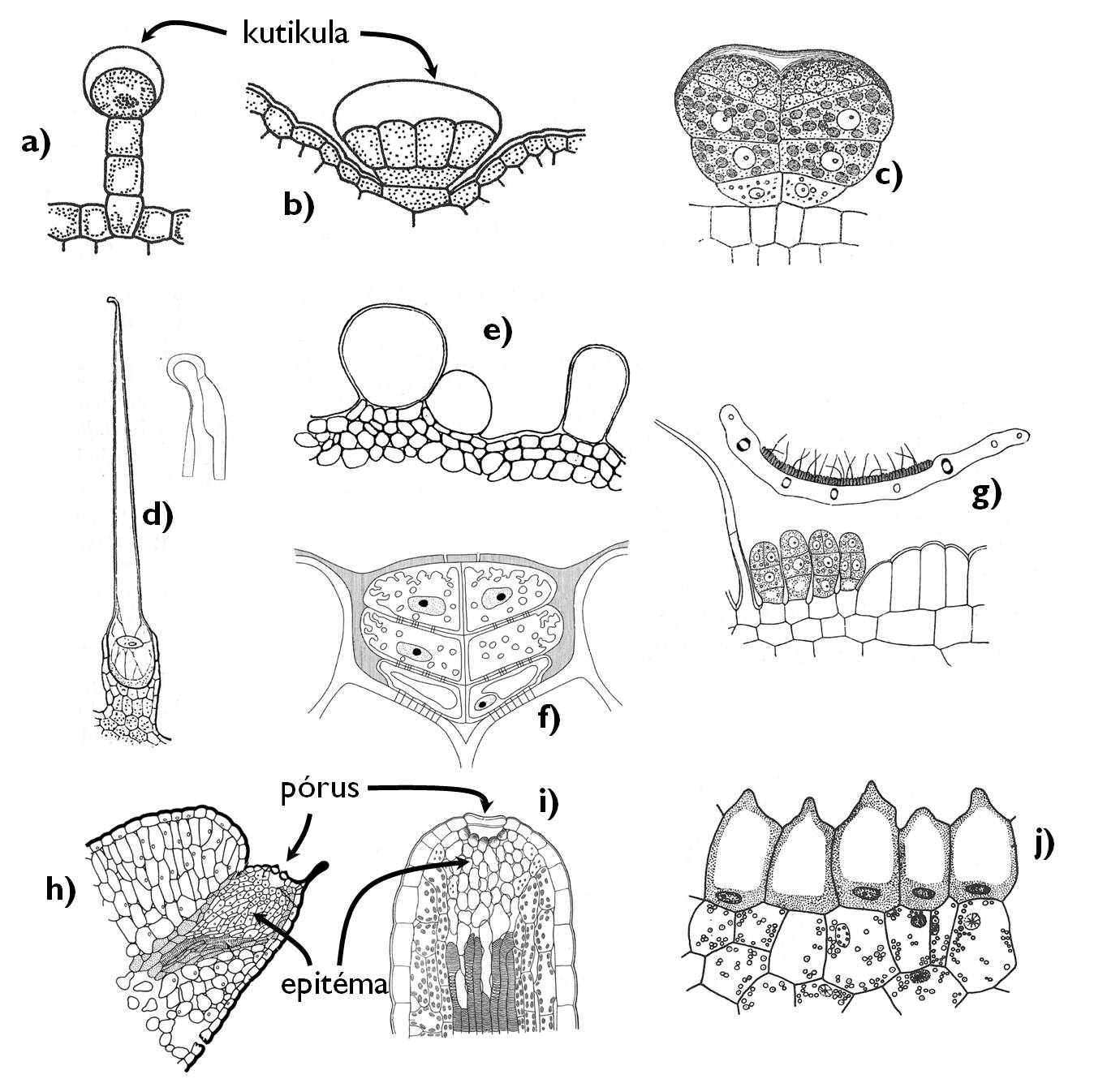
4. ábra. Az epidermális kiválasztó képletek.
a) mirigyszőr (Pelargonium); b-c) mirigypikkelyek (b: Lamiaceae, c: Asteraceae); d) csalánszőr; e) víztartó szőrök (Tetragonia tetragonoides [új-zélandi spenót]) f) sómirigyek; g) nektárium; h-i) hidatódák; j) ozmofóra.
(a-b: Jacob és mts. nyomán, c-d: Haberlandt nyomán, e: Kárpáti nyomán, f: Beck nyomán, g: Haberlandt nyomán, h: Evert és Eichhorn nyomán, i: Haberlandt nyomán, j: Haraszty nyomán)
Ha túl sok a víz: a hidatódák működése
Változatos felszíni kiválasztó képletek a magas cukortartalmú váladékot előállító nektáriumok. Két fő típusuk a virágban elhelyezkedő, a megporzók számára táplálékot biztosító intraflorális nektárium, valamint a virágon kívül (virágkocsányon, levélnyélen) elhelyezkedő extraflorális nektárium, amelyek elsősorban a növényevőkkel szemben védelmet nyújtó rovarok (többnyire hangyák) táplálására szolgálnak. Az intraflorális nektáriumok elhelyezkedése igen változatos lehet. Kialakulhatnak a vacok belső oldalán, elhelyezkedhetnek a magház körül vánkosszerűen vagy annak felső oldalát is boríthatják, de egyes virágrészek (sziromlevelek, porzók) is átalakulhatnak nektáriummá. A nektárium kiválasztószövetét alkothatja csak a módosult epidermisz (amit boríthatnak egy- vagy többsejtű mirigyszőrök is, pl. a Tilia [hárs] nemzetségben). Ennél gyakoribb, hogy egy néhány rétegből álló, alapszöveti eredetű mirigyszövet („nektarogén szövet”) termeli a cukros váladékot, ami vagy a felette levő módosult sztómákon át, vagy az epidermisz külső sejtfalának repedésein, nyílásain át jut a külvilágba. A nektár cukrok (szacharóz, glükóz és fruktóz) mellett kis mennyiségben aminosavakat, más szerves savakat, fehérjéket (főleg enzimeket), lipideket, ásványi anyagokat, foszfátokat, alkaloidokat, fenoloidokat és antioxidánsokat is tartalmazhat. Anyagai alapvetően a nektárium közelében elvégződő háncselemekből származnak, de a nedv összetétele a nektárium szöveteiben valamelyest megváltozik.
Speciális felszíni kiválasztó képletek fordulnak elő a rovarfogó és rovaremésztő növények hajtásán. A csapdaként funkcionáló módosult levelek – rendszertani csoporttól függően – sokfélék lehetnek, és az ezeken előforduló mirigyképletek is igen változatosak. Ezek három fő típusba sorolhatók: nektárt termelő csalogató mirigyek, nyálkatermelő mirigyek, amelyek váladéka csúszóssá teszi a csapda felületét (ezzel megakadályozva az áldozat menekülését) és emésztőenzimeket termelő mirigyek. Néhány faj esetében a nyálkaanyagot és az enzimeket egyazon mirigytípus termeli (pl. a Drosera [harmatfű] fajok nyeles, összetett felépítésű tentákulumai esetében). A rovarfogó fajok mirigyeinek érdekessége, hogy az anyagleadás mellett felszívást is végezhetnek. Ezeken keresztül kerülnek ugyanis a megemésztett rovartestből származó tápanyagok a növény szervezetébe.
Az epidermisz egyéb sejttípusai és feladatai
Az eddig tárgyalt sejttípusok mellett a hajtás bőrszövete további speciális sejteket is tartalmazhat, amelyek egy-egy konkrét környezeti körülményhez történő alkalmazkodást szolgálnak.
A pázsitfűfélék (Poaceae) leveleinek bőrszövetében például igen változatos sejttípusok fordulnak elő. Az epidermiszsejteket itt méretük alapján rövid- és hosszúsejtekre szokás elkülöníteni, amelyek közül a rövidsejtek speciális anyagokat tartalmazhatnak: kovasejteket (SiO2-dal merevített falú sejtek) és parasejteket (szuberinizálódott falú sejteket) találunk köztük. A száraz, meleg környezetben élő pázsitfűfélék (pl. Festuca /csenkesz/, Stipa /árvalányhaj/, Zea /kukorica/ fajok) színi levélepidermiszének jellegzetes képletei a kis csoportokban elhelyezkedő bulliform sejtek (ízületi sejtek). E sejttípus jellegzetessége, hogy a levélfelszín felé eső sejtfala jóval vékonyabb a felszínre merőlegesen futó ill. az alapszöveti sejtek felé néző sejtfalnál. Tartós szárazság esetén a növény általános vízvesztésével a bulliform sejtek víztartalma is lecsökken. A turgorcsökkenés hatására – az egyenlőtlen sejtfalaknak köszönhetően – a bulliform sejtek külső felülete összeesik, ezáltal meghajtják a levélfelületet. Mivel több ilyen sejtcsoport is előfordul a levél felületén, a meghajlás összeadódik: a vízvesztés következtében a teljes levél a hossztengelye mentén besodródik (a kukorica esetében erre a jelenségre mondják, hogy „furulyázik” a növény). A folyamat előnye, hogy a levélhenger belsejében a párologtatás miatt páratelt tér jön létre, ami csökkenti a növény további vízleadását, így segítve a száraz környezethez történő alkalmazkodást.
Egyes növényfajok levelének epidermisze vízraktározásra képes, a víztartó alapszövethez (lásd később) hasonlóan olyan sejtekből épül fel, amelyekben nagyméretű, nyálkaanyaggal telt vakuólum található. A poliszacharid (vagy glikopoliszacharid) természetű nyálka a víztartó alapszövethez hasonlóan biztosítja a sejt víztároló képességét. Ilyen epidermisszel találkozunk a Commelinaceae /kommelinafélék/ családjában (pl. Zebrina pendula /zebrapletyka/).
Az epidermisz anatómiai jellemzői a fénymennyiséghez történő alkalmazkodást is szolgálhatják. Az árnyéktűrő növények levelének fonáki bőrszövete gyakran a vakuolumban felhalmozódó antociánoktól lilás színű (pl. Zebrina pendula /zebrapletyka/, Cyclamen purpurascens /erdei ciklámen/). A lila színű festékanyag ebben az esetben arra szolgál, hogy a levél mezofillumán áthaladó, de a fotoszintézisben nem hasznosult fény egy részét visszaverje a kloroplasztiszok irányába, ahol esély van arra, hogy ismét részt vegyen a fotoszintézisben.
A másodlagos bőrszövet: a periderma és a ritidóma
Abban az esetben, ha a növényi szervek növekedése az elsődleges szövetek kialakulása után is folytatódik, másodlagos növekedésről beszélünk. A szervek másodlagos növekedését a szerveket borító elsődleges bőrszövet (rizodermisz illetve epidermisz) – a sejtosztódás hiánya miatt – nem képes követni, ezért leszakadozik. Természetesen ezek a szervek sem maradhatnak védelem nélkül: felületükön másodlagos merisztéma eredetű védőréteg, a másodlagos bőrszövet (periderma) alakul ki.
A másodlagosan vastagodó szár, gyökér (néha a termés) epidermiszének leszakadozása után az alatta elhelyezkedő szövetréteg (alapszövet) kerül a szerv felszínére. Ennek meghatározott sejtjei visszanyerik osztódóképességüket (dedifferenciálódnak) és másodlagos merisztémává, a parakambiummá (fellogén) alakulnak. (A gyökérben nem a legkülső sejtsor alakul fellogénné, hanem a központi henger legkülső sejtsora, a periciklus, valamint az azt körülvevő alapszöveti sejtek együttesen hozzák létre a fellogént.) A parakambium sejtjeinek osztódásai a külvilág felé (monopleurikus fellogén) vagy mind a külvilág, mind pedig a szerv belső szövetei felé (bipleurikus fellogén) új sejteket hoznak létre. A külvilág felé eső szövetréteg a paraszövet (fellom), a szerv belseje felé néző (egyes esetekben hiányzó) réteg a paraalapszövet (felloderma). A periderma tehát a fellom, a fellogén és – ha van – a felloderma együttese.
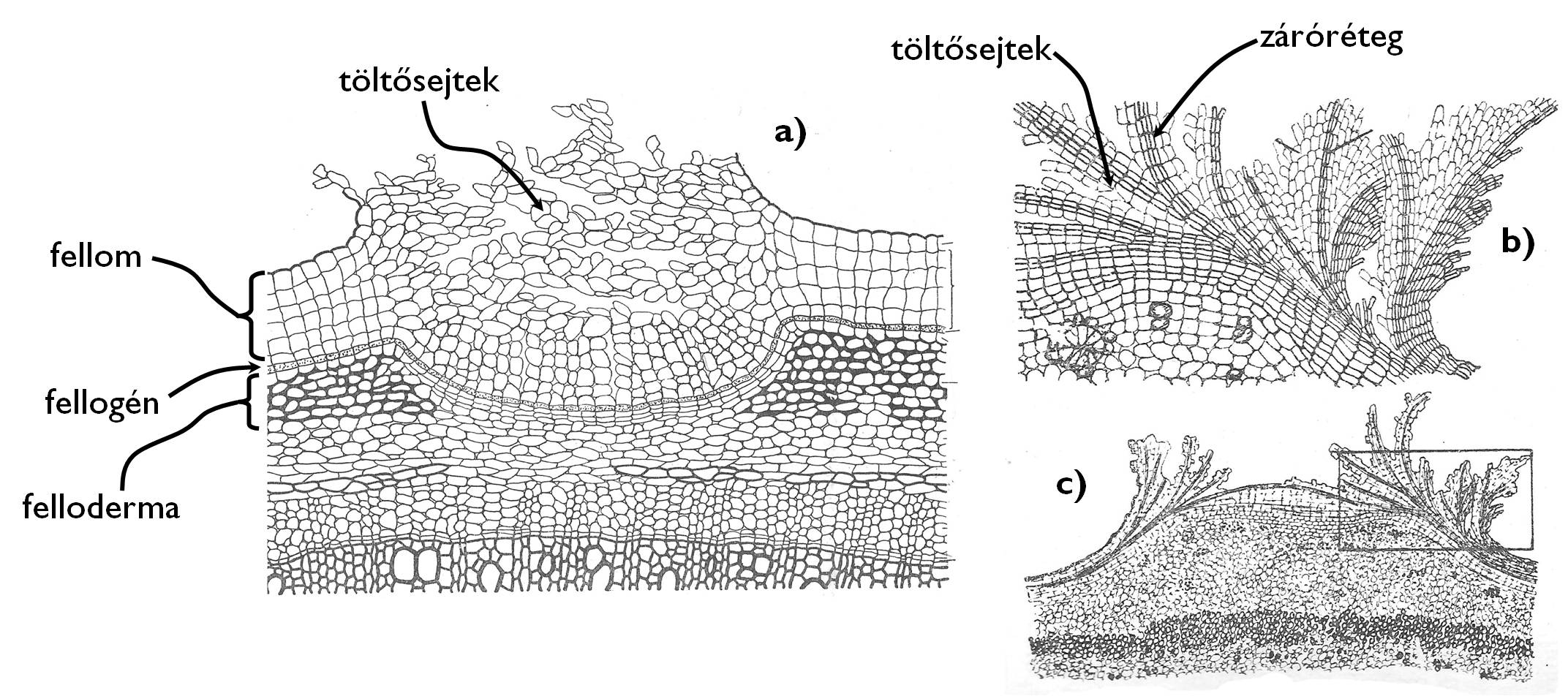
5. ábra. A periderma és a lenticella feléptése.
a) Sambucus-típusú lenticella, b-c) Gleditsia-típusú lenticella.
(Sárkány és Szalai nyomán)
A periderma magyar neve („parabőr”) arra utal, hogy sejtrétegeit szuberinizálódott falú sejtek alkotják. A paraanyag berakódásának köszönhetően ezek a sejtek nem engedik át a vizet, viszont maguk a sejtek elhalnak (természetesen a fellogén sejtjeinek kivételével). A periderma sejtrétegei azonban nem csak a vizet, de mivel gázcserenyílásokat nem tartalmaznak, a levegőt sem eresztik át, Annak érdekében, hogy a másodlagos bőrszövettel fedett szervek szövetei is hozzájussanak a légzési gázokhoz, azokon a helyeken, ahol az epidermiszen eredetileg a sztómák helyezkedtek el, kráterszerű nyílások, az ún. paraszemölcsök (lenticella) jönnek létre (5. ábra). A paraszemölcsök területén a periderma felső rétege (a fellom) felszakad és elemelkedik az alsóbb rétegektől. A fellom alatti területet speciális, a levegő számára átjárható, sejt közötti járatokkal átszőtt alapszövet tölti ki, amit szintén a parakambium hoz létre. (Lényegében a paraszemölcs a fokozott fellogén-működés miatt kialakuló periderma-burjánzás, ahol a gyors osztódás miatt a sejtek eltávolodnak egymástól.)
A töltőszövet változatossága: a lenticellák típusai
A fás szárú növények hajtásai több éven át folyamatosan vastagszanak, így peridermájuk folytonosan tágul, felrepedezik. Ezzel párhuzamosan a másodlagos bőrszövet alatti alapszövetnek újabb és újabb sejtsora alakul fellogénné. Ez azonban azzal jár, hogy a szállítószöveteket körülvevő alapszövetréteg (az elsődleges kéreg – lásd a szár szövettanánál!) folyamatosan fogy. Amikortól a szállítószövet háncsrésze felett már nincs olyan alapszövet, amelynek dedifferenciálódása a parakambium kialakulását biztosíthatná, a háncstest legkülső alapszöveti elemei hozzák létre a fellogént. A háncstestből létrejövő osztódószövet létrehozta periderma ezért már különböző háncselemeket (rostasejtek, rostacsövek maradványait, háncsrostokat) tartalmaz. Ez a floem-eredetű védőszövet a héjkéreg (ritidóma). Makroszkópos megjelenése fajra jellemző, a háncstest és a parásodott sejtek szerkezetétől függően különböző színe, megjelenése, leválási módja határozóbélyeg a fás szárú növények esetében.
A szállítószövet-rendszer
Anyagszállítási útvonalak a növényi szervezetben
A növényi szervezetben a különféle anyagok sejtről sejtre történő szállítása kétféle útvonalon valósulhat meg (6. ábra). Az egyik az ún.apoplasztikus szállítás, amikor a sejtek protoplazmáján (azaz a sejthártyán) kívüli térben levő vizes oldatban mozognak a vegyületek. Az apoplasztot a sejtfal üregrendszerei és a sejtek közti (intercelluláris) járatrendszer alkotja. A másik útvonal a sejtek plazmodezmákkal összekapcsolt protoplazma-rendszere, a szimplaszt. A szimplaszt valójában a plazmalemmán belül elhelyezkedő területek összességét jelöli. A plazmodezmáknak köszönhetően (1ásd az 1. fejezetben!) az egész növényi szervezet néhány nagyobb szimplasztikus egységből áll.
Az apoplasztból a szimplasztba történő átlépés a sejthártyán keresztül történhet meg, apoláris anyagok esetében diffúzióval, máskor valamilyen fehérje közvetítette transzport révén (részletesen lásd a sejttani részben leírtakat). A két térrészben zajló szállítás közti legfontosabb különbség, hogy a szimplasztban zajló transzport szabályozható. Éppen ezért mindig jelentősége van annak, ha valamilyen anyag szállításának útvonala a szimplasztba helyeződik át (pl. a gyökérben az endodermisz sejtsorában történik ilyen – részletesen lásd a gyökérnél).
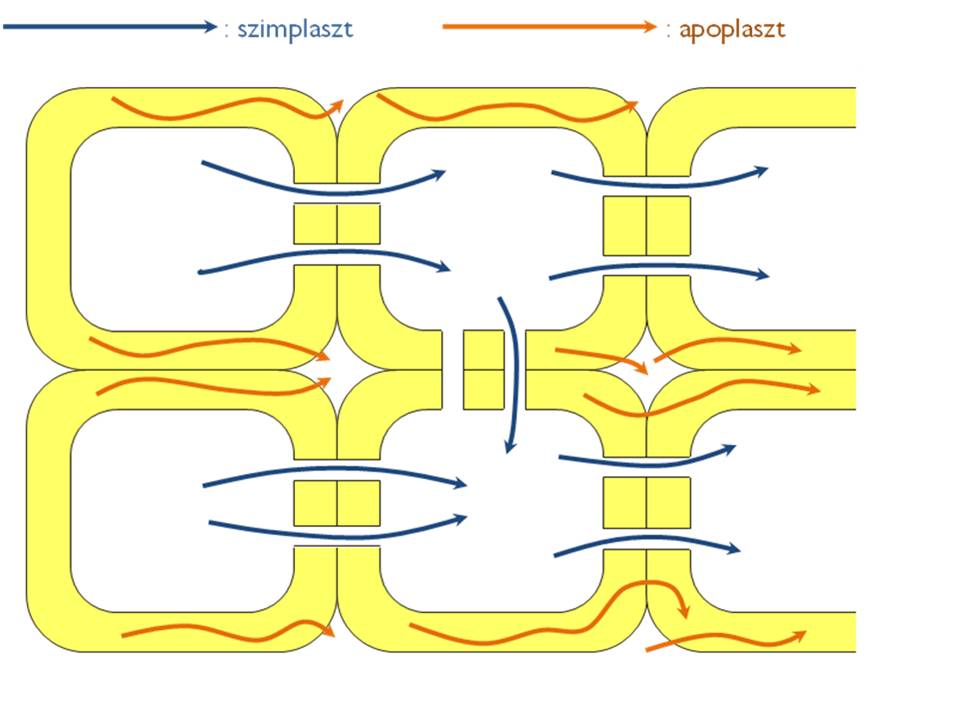
6. ábra. Az anyagszállítás útvonalai a növényi testben.
A növényi szervezetben szállított anyagokat két nagy csoportra oszthatjuk. A tápanyagok, azaz növények esetében a víz és az ásványi anyagok (szervetlen ionok) a gyökér felől haladnak a többi szerv (lombozat, zöld színű szárak, érőben levő gyümölcsök) felé. A szerves anyagok szállítása mindig termelődésük helye felől, a felhasználás irányába történik. A fotoszintézissel létrehozott cukor (szállított formája a szacharóz) és az aminosavak esetében a szállítás iránya a kifejlett levelek felől a fejlődő növényi részek illetve a raktározó szervek irányába mutat. A növényi hormonok mozgása az előállító sejtek felől a célszövetek felé történik.
A növényi testben zajló hosszú távú, irányított (polarizált) transzport fő színtere a szállítószövet-rendszer, illetve az abból felépülő szállítónyaláb. Emellett azonban rövid távú, közeli sejtek között zajló szállítás is folyik a növényi szervezetben, amit alapszöveti sejtek (a parenchimasejtek, pl. bélsugarak vagy ún. transzfersejtek – lásd később) végeznek. A rövidtávú szállítás nem igényel speciális alaki alkalmazkodást a sejtektől, a hatékony transzportot megfelelő rendeződésük (pl. a bélsugarakban) vagy az intenzív sejt-sejt kapcsolatokat lehetővé tevő nagyobb sejtfelszín (a transzfersejtek esetében) szolgálja. Ezzel szemben a szállítószövet-rendszerben levő sejtek a hatékony szállítás érdekében megnyúlnak, átmérőjük megnő (csőszerűvé válnak), protoplazmájuk kisebb-nagyobb mértékben degenerálódik az anyagszállítás akadálytalanná válása céljából. A hosszúra nyúlt sejtek merevítését jellegzetes módon (spirálisan, gyűrűsen) vastagodott másodlagos sejtfal szolgálja. E szövetrendszer tagjai kloroplasztiszokat nem tartalmaznak, közöttük- a bőrszövetrendszerhez hasonlóan – kevés a sejt közötti állomány.
A szállítószövet-rendszer gyakran kapcsolódik alapszöveti elemekkel: a szállítónyalábok felépítésében mindig részt vesznek parenchimasejtek (fa- ill. háncsparenchima) és szilárdító rostok (fa- ill. háncsrostok) is.
A szállítószövet-rendszer felépítése
A szállítószövet-rendszer elemeit funkciójuk alapján két csoportba sorolhatjuk, amelyek a térben is elkülönülten helyezkednek el. A farész (xilem) feladata a növényi tápanyagok (víz és ásványi sók) szállítása a gyökértől a lombozat irányába, míg a háncsrész (floem) végzi a szervesanyag-szállítást (cukrok, aminosavak) a termelés helye felől a felhasználás irányába.
Fejlődési állapotuk alapján többféle fa- és háncselemet különböztetünk meg. Az elsőként megjelenő szállítóelemeket a megfelelő elsődleges merisztémák (a szárban a prokambium, a gyökérben a pleróma) osztódó sejtjei hozzák létre: ezek az elsődleges (primer) xilem és az elsődleges floem sejtjei.
Az elsődleges szállítószövet változatossága: proto- és metaelemek
A másodlagosan vastagodó szervekben a differenciálódott szövetek másodlagos merisztémákká alakulhatnak, amelyek közül a különböző kambiumtípusok (vaszkuláris kambium, hullámos kambium) szállítóelemeket is létrehoznak. Az eképpen létrejövő szállítószövet a másodlagos (szekunder) xilem és a másodlagos floem.
A farész (xilem) felépítése és működése
Az ásványi anyagok és a víz szállítását végző farész tagjai a szállítóelemek (vazális elemek), a faparenchimasejtek (mindezek összessége az ún. hadrom) valamint a szilárdító feladatú farostok (xiláris rostok) (1. táblázat).
A farészben a szállítás mindig egyirányú (a gyökértől a lombozat felé folyik) s tisztán fizikai-kémiai törvényszerűségeken alapul. A gyökérszőrsejtek folyamatos ozmotikus vízfelvétele nyomóerőt gyakorol a farészben haladó vizes oldatra (gyökérnyomás), aminek a mozgását a farész elemeiben fellépő kapilláris-szívóerő is segíti. A vízszállításban részt vevő harmadik mozgató erő a párologtatás szívóereje, ami azért jön létre, mert a párologtatást végző szervekben a szövetek sejt közötti állományában jóval nagyobb a vízmolekulák koncentrációja, mint a külvilágban, ezért azok diffúzióval folyamatosan távoznak a légtérbe. A felsorolt hajtóerők kialakításához nincs szükség élő sejtekre, ezért a hatékony szállítás érdekében a faelemek protoplazmája a differenciálódás során felszívódik (a sejtek apoptózissal elpusztulnak). Ezért a xilem sejtjei – a faparenchimasejtek kivételével – holt sejtek.
Valamennyi növénycsoport farészében találunk vízszállító sejteket (tracheidákat) (7. ábra). Ezek megnyúlt, lignifikált falú sejtek, amelyek kihegyesedő végeikkel sorakoznak egymás fölött. Az átfedő sejtvégeken gödörkésen (vagy nyitvatermők esetében vermes gödörkésen) vastagszik a sejtfal, az oldalsó részeken többnyire gyűrűsen vagy spirálisan. A szállítás a haránt irányú sejtfalakon levő gödörkepárokon keresztül történik: a szállítószövet hossztengelyéhez képest tehát egy emelkedő spirál mentén.
A zárvatermőkben a tracheidák mellett megjelennek a vízszállító csövek (tracheák) is. Ezek az igen hosszú (akár több méteres) elemek viszonylag rövid, de igen nagy átmérőjű sejtek (tracheatagok) összeolvadásával jönnek létre, a haránt irányú sejtfalak fokozatos felszívódása révén. A tracheák igen nagy átmérőjű elemek, ezért a vízszállító sejteknél gyorsabb, hatékonyabb, tengelyirányú szállítást tesznek lehetővé. Megjegyzendő azonban, hogy a tracheák sokkal érzékenyebbek a légbuborékok (embóliák) okozta eltömődésre, mivel a bennük kialakuló buborékok összegyűlhetnek, és nagyobb szakaszokat zárhatnak el, mint a tracheidákban. A vízszállító sejtek esetében ezzel szemben a kialakuló buborékok a tracheida csúcsában megrekednek, ami nem akadályozza a harántfalakon át történő vízszállítást. Ez a magyarázata annak, hogy a nyitvatermők körében miért lehet olyan sok örökzöld, tehát a téli, fagyos időszakban is vízszállítást végző faj. A mérsékelt és hideg övben élő zárvatermők ezzel szemben a téli időszakban lehullatják a lombjukat, így védekezve a téli fagyokat követő olvadás során hirtelen kiváló légbuborékok tracheákat elzáró hatásával szemben. (A téli nyugalmi időszak ellenére a zárvatermők farészében sokkal rövidebb ideig működőképesek a xilem-elemek, mint a nyitvatermők tracheidái.)
A vazális elemek mellett a farészben mindig találunk alapszöveti sejteket is; ezek a faparenchimasejtek. Feladatuk a raktározás illetve a rövid távú szállítás. Két rendszerben fordulnak elő a xilemben: egyrészt a tracheidákkal, trancheákkal párhuzamosan rendeződve (szűk értelemben csak ezeket szokták faparenchimasejteknek nevezni), másrészt ezekre merőlegesen, a bélsugarakat alkotva. Az utóbbiak feladata a haránt irányú szállítás és a fatest rugalmasságának biztosítása is.
A zárvatermők farészének szilárdító elemei a hosszú, kis átmérőjű, szűk üregű farostok. E szilárdító alapszöveti sejtek erősen lignifikálódott faluknak köszönhetően fokozzák a vazális elemek mechanikai stabilitását. A nyitvatermők farészében nem találunk farostokat, szerepüket vastag falú, a szállítás helyett első sorban szilárdító funkciót betöltő rosttracheidák látják el. (Ezekből több, átmeneti forma fordul elő a nyitvatermők fájában, a szállítást is végző, de vastagodott falú tracheidáktól kezdve a csak csökevényes vermes gödörkéket hordozó, szállításra képtelen típusokig.)
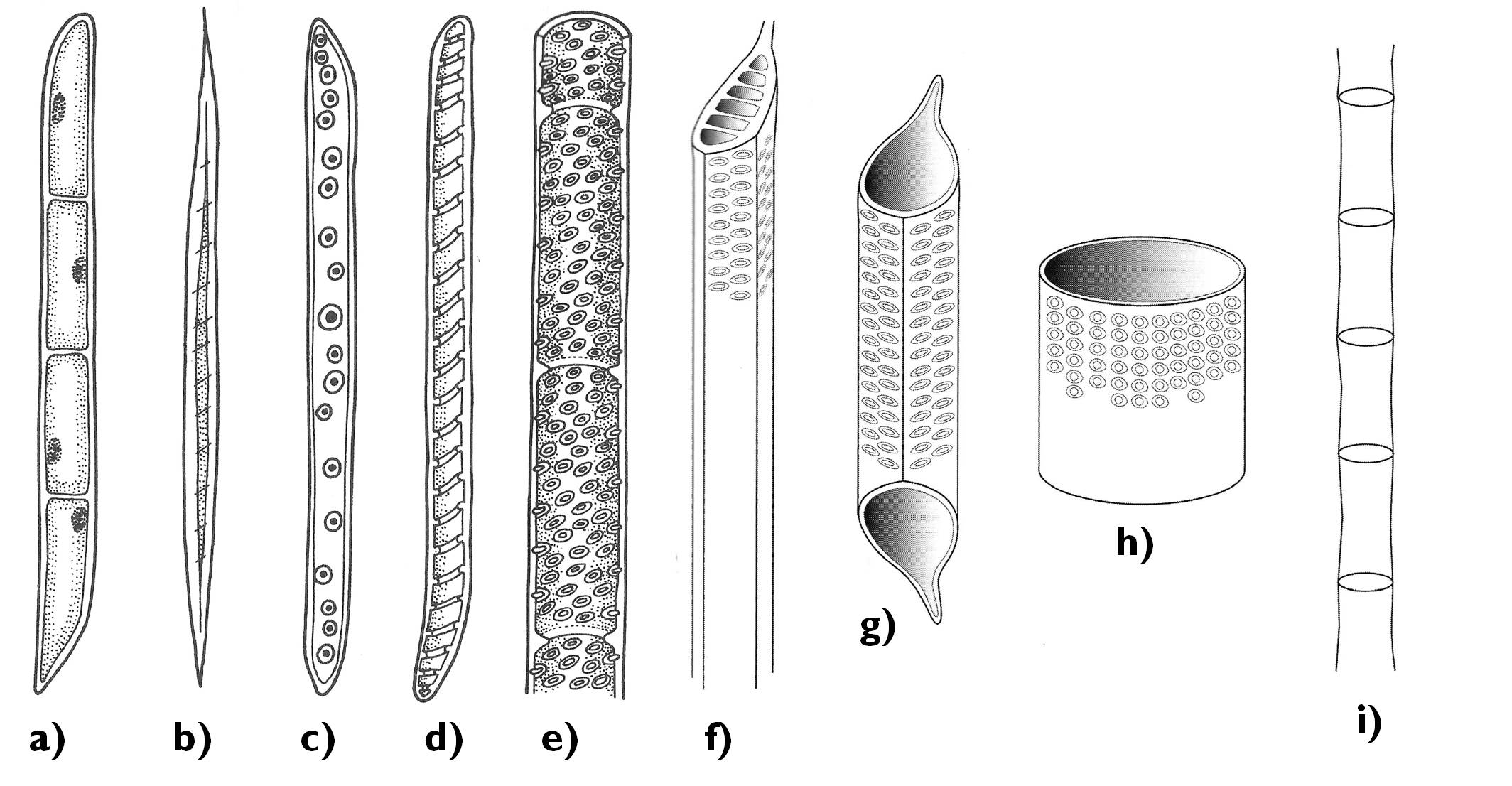
7. ábra. A xylemelemek típusai.
a) faparenchima-sejtek; b) farost; c-f) tracheidák (c: vermes gödörkés sejtfal, d: gyűrűs sejtfal, e-f: gödörkés sejtfal); g-h) tracheatagok; i) trachea.
(a-e: Jacob és mts. nyomán, f-i: Beck nyomán)
A háncsrész (floem) felépítése és működése
A háncsrész (floem) feladata a fotoszintézis során termelt asszimilátumok répacukor (szacharóz) formájában történő szállítása. Emellett aminosavak és egyes hormonok szállítása is itt zajlik. A háncsrész szállításban részt vevő elemei (kribrális elemek) az alapszöveti háncsparenchimasejtek segítségével látják el feladatukat (összességük a leptom). A floem szilárdságát a háncsrostok (extraxiláris rostok) biztosítják (1. táblázat).
1. táblázat. A szállítóelemek rendszere.
-
floem
háncsrostok
leptom
háncsparenchima-sejtek
kribrális elemek
rostasejtek
rostacsövek
kísérősejtek
xilem
farostok
hadrom
faparenchima-sejtek
vazális elemek
tracheidák
tracheák
A szervesanyagok szállítása a termelés helye felől („forrás” szervek) a felhasználás („nyelő” szervek) irányába történik. Ennek megfelelően a növényben vannak olyan háncselemek (pl. a levélnyelekben), amelyeken keresztül változik a szállítás iránya. A háncsrészben tehát szabályozott transzportfolyamat zajlik, amihez protoplazmával rendelkező sejtek kellenek, a floem tagjai tehát – a rostok kivételével – élő sejtek. A kribrális elemek esetében azonban a sejtplazma összetétele megváltozik: a differenciálódás során a sejtnedv-vakuolum membránja felbomlik és a vakuolum tartalma összekeveredik a sejtplazmával (ún. mixoplazma jön létre), majd egyes sejtalkotók (pl. a sejtmag) is lebomlanak.
A háncsrész legkisebb méretű szállítóelemei a harasztokra és nyitvatermőkre kizárólagosan jellemző rostasejtek (8. ábra). Ezek a megnyúlt, spirálisan, gyűrűsen vastagodott falú sejtek rézsútosan futó, átfedő sejtfalaikon átnyúló plazmodezmáik révén adják át egymásnak a szervesanyagokat. A plazmahidaknak köszönhetően szitaszerűen áttört sejtfalfoltok a rostamezők.
A zárvatermők floemjében a rostasejtek mellett megjelennek a rostacsövek. Ezek – a tracheákhoz hasonlóan – rövidebb, de nagy átmérőjű egységekből, a rostacsőtagokból állnak. A rostacsőtagok között az érintkező harántfalból a plazmodezmák kiszélesedésével létrejövő, sűrűn átlyuggatott rostalemez tart kapcsolatot. A rostalemezt áttörő plazmafonalak köré kallóz rakódik, ami később az egész haránt sejtfalat beborítja, míg végül – a rostacsövek működésének megszűnésekor – a lyukakat is eltömi.
A rostacsövek és a rostasejtek anyagcseréjét és működését a velük szomszédos sejtek segítik. A rostacsövek keletkezése során a rostacső-anyasejt inekvális osztódásával a rostacsőtag mellett egy másik, keskenyebb sejt, a kísérősejt jön létre, ami számos plazmodezma révén a differenciálódás után is megőrzi szoros kapcsolatát a rostacsőtaggal. A kísérősejtek sűrű citoplazmájú, aktív anyagcserét folytató sejtek, különféle anyagokat adnak át a rostacsőtagnak (illetve vesznek fel belőle). Szoros kapcsolatukra utal a tény, hogy az egyik pusztulása a másik elhalását okozza. (+info)
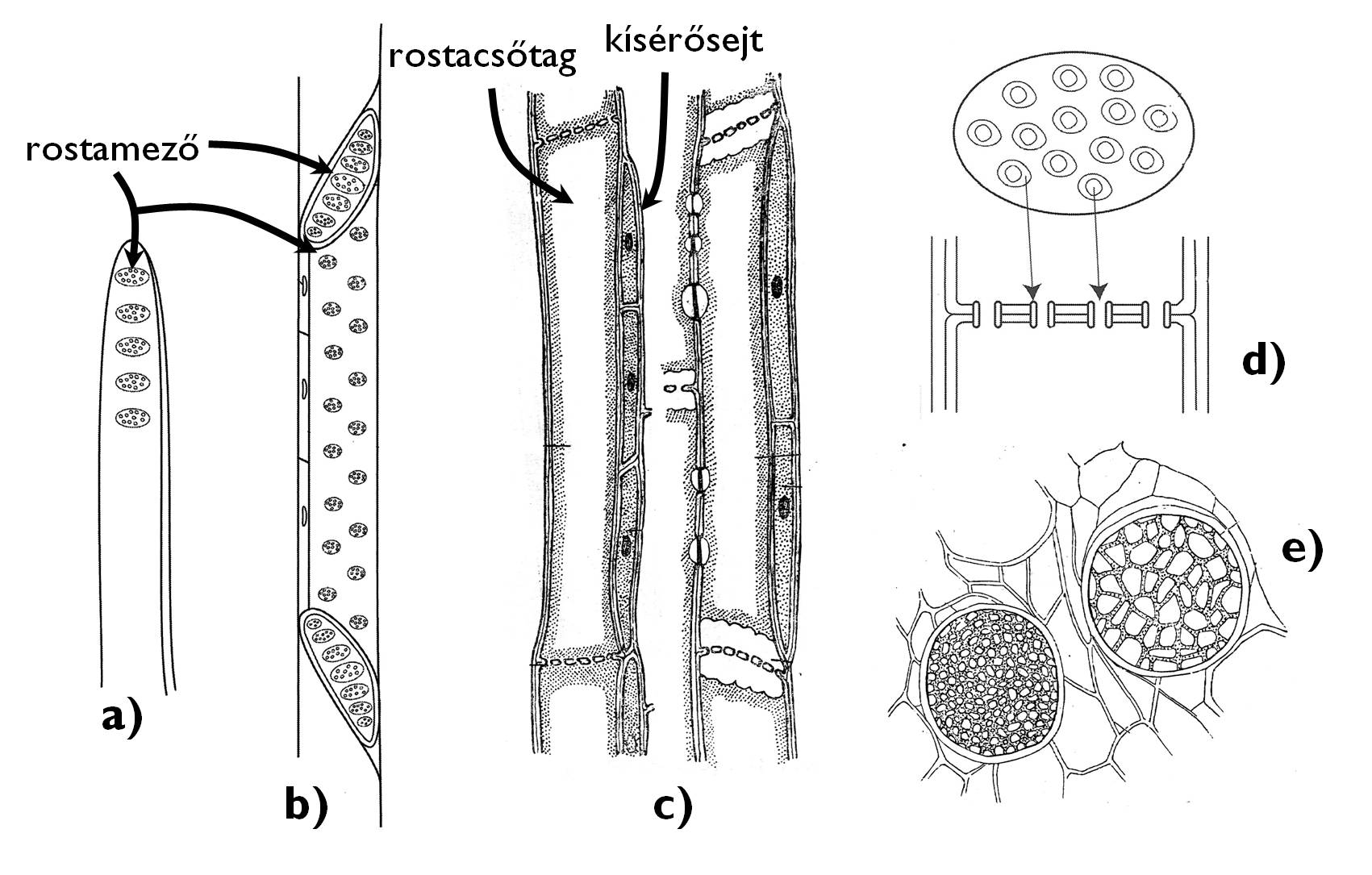
8. ábra. A phloemelemek típusai.
a-b) rostasejt; c) rostacsőtag kísérősejtekkel; d-e) a rostalemez szerkezete.
(a-d: Beck nyomán, e: Habelrandt nyomán)
A háncsrészben szállított szénhidrátok esetében a forrásszerv többnyire a kifejlett, fotoszintetizáló levél (esetleg a zöld színű szárak), míg nyelők lehetnek a raktározó szervek (gumó, rizóma stb.), a növekvőben levő részek (fiatal levelek, hajtás-, gyökércsúcs) vagy a fejlődőben levő virágok, termések, magvak. Ennek megfelelően a fiatal levelek nyelében található háncselemekben eleinte a szár felől a fejlődő levéllemez irányába mutat a szállítás, míg a levél kialakulását (és az aktív fotoszintézis megindulását) követően a transzport iránya megfordul: ekkor már a levélalap (azaz a szár) irányába mutat.
Szállítás a floemben: a tömegáramlás-hipotézis
A szállítónyalábok
A növényi szervezetben a szállítószövet elemeit soha nem elszórva, magányosan találjuk; azok kötegeket, szállítónyalábokat alkotnak. A szállítónyalábok felépítésében a vazális illetve a kribrális elemek mellett alapszöveti (parenchimatikus és szilárdító) sejtek is részt vesznek.
A szállítónyalábokat a bennük előforduló háncs- és faelemek megléte, valamint azok kölcsönös térbeli helyzete alapján csoportosíthatjuk (9. ábra). Az egyszerű szállítónyalábokban vagy csak fa- vagy kizárólag háncselemek fordulnak elő. Ezt a nyalábtípust a gyökér elsődleges szöveti szerkezetében figyelhetjük meg. A hajtásban (azaz a szárban, a levélben és a generatív szervekben) ezzel szemben mindig xilemet és floemet egyaránt tartalmazó, ún. összetett nyalábokat figyelhetünk meg. Az összetett nyalábokban a fa- és a háncselemek egymáshoz képest többféleképpen helyezkedhetnek el. A kollaterális szállítónyalábban a xilem és a floem egymás mellett fut, egy közös határfelületen érintkezve egymással, míg a koncentrikus nyalábokban valamelyik szállítószövet típus teljesen körülveszi a másikat. A lemezes nyaláb esetében a háncselemek lapos kötegekben, lemezekben váltakoznak a xilem elemeivel (ezt figyelhetjük meg pl. a Lycopodium clavatum [kapcsos korpafű] kúszó hajtásaiban).
A kollaterális nyalábok fa- és háncsrésze közé osztódó szövet (kambium) ékelődik abban az esetben, ha az vastagodni képes szervben (pl. örökzöld levélben vagy valódi kétszikű /Rosopsida/ növény vastagodó szárában) helyezkedik el; ezt a nyalábtípust nevezzük kollaterális nyílt nyalábnak. Amennyiben a xilem és a floem között nem találunk kambiumot (pl. a további vastagodásra nem képes lágyszárakban vagy a levelek többségében), kollaterális zárt nyalábról beszélünk. Egyes családok (pl. Apocynaceae [meténgfélék], Cucurbitaceae [tökfélék]) kollaterális nyalábjaiban a farészt két oldalról (kívülről és belülről) is floem-elemek fogják közre (de nem veszik azt teljesen körül); ez a bikollaterális nyaláb (+info).
A koncentrikus nyalábokat aszerint csoportosíthatjuk, hogy a háncsrész vagy a farész halad-e a nyaláb közepén. Amennyiben a háncsrész veszi körül a faelemeket amfikribrális (más néven hadrocentrikus) nyalábról beszélünk. Ez a típus a páfrányok rizómájában (pl. a Pteridium aquilinum [saspáfrány] fajnál) gyakori. Az amfivazális (leptocentrikus) nyaláb középső régiójában a háncselemek helyezkednek el és ezeket övezi gyűrűszerűen a xilem. (A Convallaria majalis [májusi gyöngyvirág] tarackjaiban találunk például ilyen nyalábokat.)
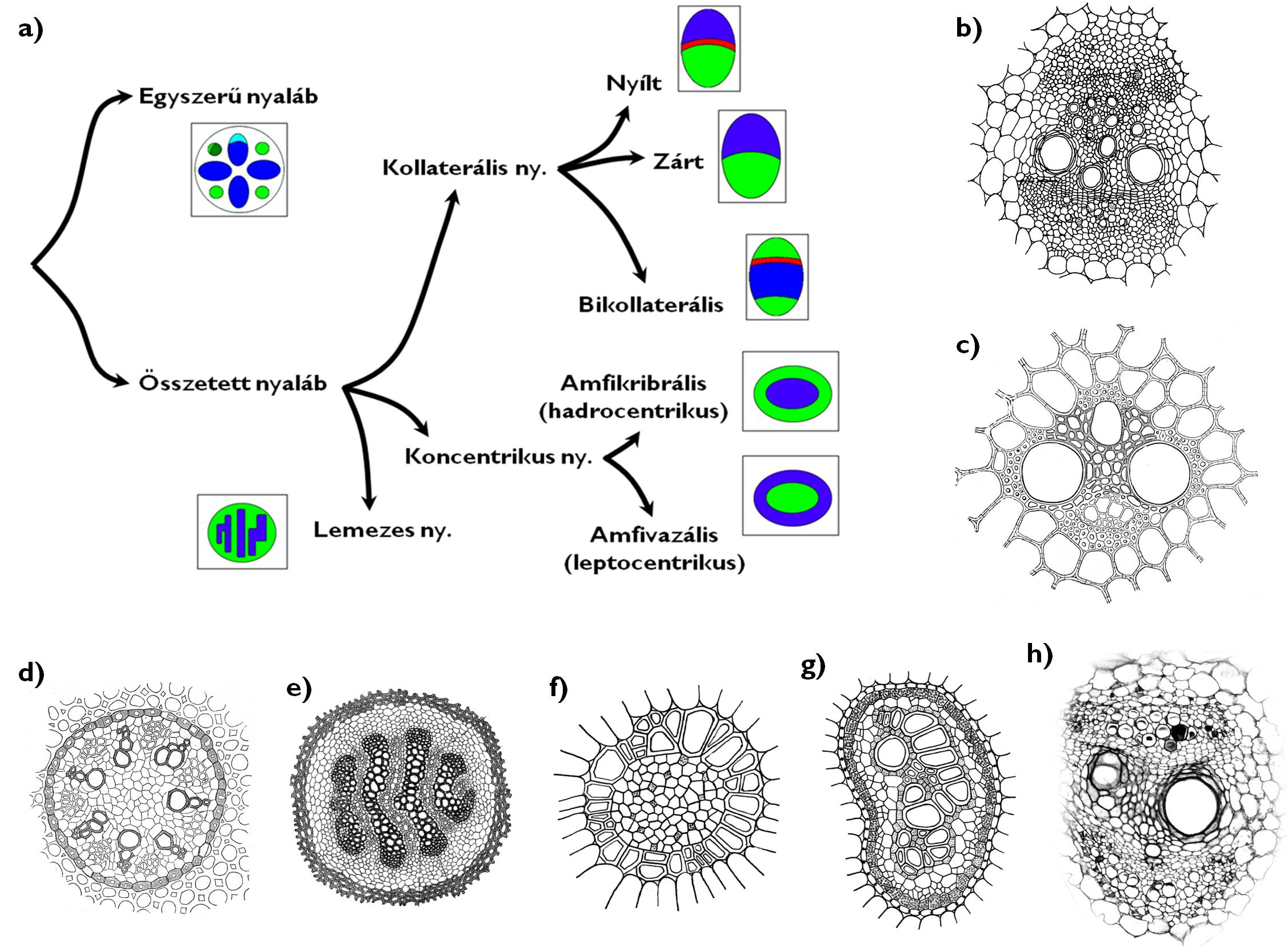
9. ábra. A nyalábtípusok.
a) a nyalábtípusok rendszere (a zöld szín a háncsrészt, a kék szín a farészt, a piros sáv a kambiumot jelöli); b) kollaterális nyílt nyaláb; c) kollaterális zárt nyaláb; d) egyszerű nyalábok; e) lemezes nyaláb; f) amfivazális (leptocentrikus) nyaláb; g) amfikribrális (hadrocentrikus) nyaláb; h) bikollaterális nyaláb.
(b: Haraszty nyomán, c-d: Habelrandt nyomán, e: Sárkány és Szalai nyomán, f-g: Haraszty nyomán, h: Evert és Eichhorn nyomán)
Az alapszövetrendszer
A növényi szervezet legáltalánosabb állandósult szövettípusa a szervek alapállományát képező alapszövet. Sejtjei igen változatos funkciókat láthatnak el, ennek megfelelően az alapszöveti sejteknek nincsen egységes morfológiai megjelenése, hiszen szerkezetük a szövet szerepének megfelelően változik. Általános jellegzetességük azonban, hogy bennük sejtközötti járatok (intercellulárisok) nyílhatnak. E szövetrendszer tagjai között találjuk a legkevésbé differenciálódott szöveti elemeket is, amelyek a leginkább hajlamosak visszanyerni osztódóképességüket, azaz belőlük jönnek létre leggyakrabban másodlagos merisztémák.
A sejtközötti terek változatossága
Az alapszövetek típusai
Az alapszövetek legáltalánosabb típusa a valódi alapszövet (parenchima) (10. ábra). E szövettípus sejtjei izodiametrikusak, erősen vakuolizáltak, másodlagos sejtfaluk vékony. Színtesteket többnyire nem tartalmaznak (esetleg proplasztiszok vannak bennük), köztük több-kevesebb intercelluláris fordul elő. Általános térkitöltő szerepe van – a szár, gyökér bélszöveteként vagy nem fotoszintetizáló levelek mezofillumában figyelhetjük meg.
Vékony falú, kloroplasztiszokban gazdag alapszövet a fotoszintetizáló alapszövet (klorenchima). Zöld színű növényi részekben fordul elő: levelek mezofillumában, fiatal szárak kortexében, éretlen termések falában. Sejtjei változatos alakúak: a paliszád klorenchimát oszlop alakú, szorosan záródó sejtek alkotják, a szivacsos klorenchima sejtjei között tág skizogén intercellulárisok vannak, míg a nyitvatermők tűleveleire jellemző karos paliszád klorenchima sejtjeinek belsejébe a sejtfal karszerű nyúlványai türemkednek be.
Leukoplasztiszokban gazdag, tartaléktápanyagokat felhalmozó alapszövet a raktározó alapszövet, amelyet különböző akkumuláló szervekben (karógyökér, rizóma kortexében, ággumóban) figyelhetünk meg. Altípusának tekinthető a szukkulens (pozsgás) növényekre jellemző víztartó alapszövet, amelynek sejtjeiben óriási vakuólum tárolja a vizet a benne felhalmozódó, higroszkópos tulajdonságú poliszacharid (nyálkaanyag) segítségével. Az átszellőztető alapszövet (aerenchima) a vízi növények szerveiben vagy állandó vízborítású talajban levő növényi részek levegőtől elzárt részeihez juttatja el a levegőt kiterjedt intercelluláris járatrendszere segítségével.
Szállítás az alapszövetben: a transzfersejtek
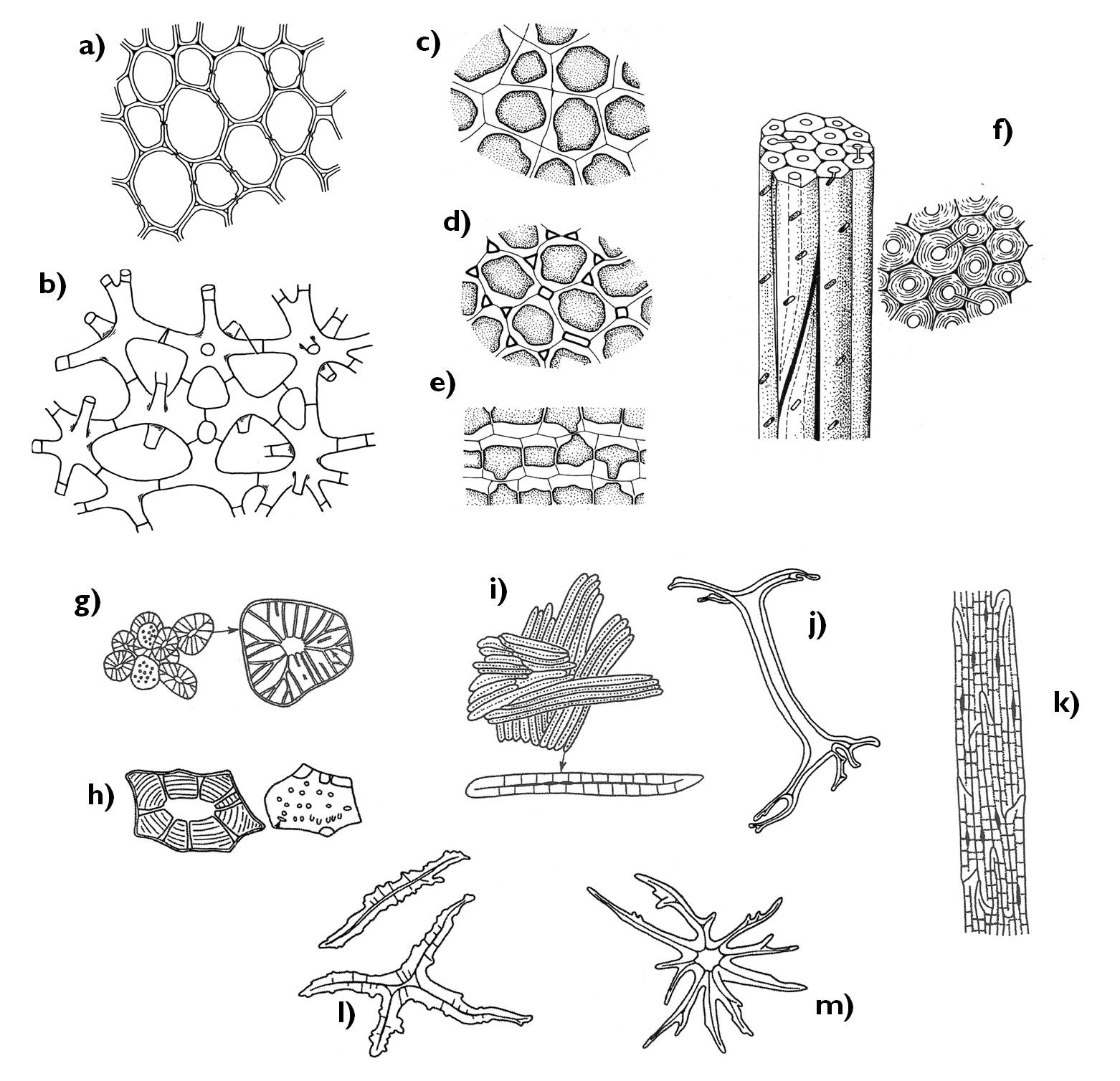
10. ábra. Az alapszövetek típusai.
a) parenchima; b) aerenchima; c-e) kollenchima (c: gyűrűs kollenchima, d: hézagos kollenchima, e: lemezes kollenchima); f) szklerenchima-rostok; g-m) szklereidák (g-h: kősejtek (brachiszklereidák) (Pyrus, Hoya), i-k: makroszklereidák (Malus, Hakea, Allium), l-m: aszteroszklereidák (Camellia, Trochodendron).
(a-b: Evert és Eichhorn nyomán, c-f: Jacob és mts. nyomán, g-m: Beck nyomán)
A szilárdító alapszövetek feladata a növényi szervek tartásának, merevségének biztosítása, valamint az érzékenyebb képletek védelme. Sejtjeik minden esetben vastag másodlagos sejtfallal rendelkeznek, ám a másodlagos sejtfal anyagai eltérőek lehetnek. A kollenchima esetében cellulózból, hemicellulózból és pektinből álló rétegek rakódnak az elsődleges sejtfalra.
A kollenchima elhelyezkedése elsősorban hipodermális (epidermisz alatti). Ez a szövettípus merevíti a fiatal szárak kérgi régióját, a leveleket a levélerek közelében, a virágok és termések kocsányát, de találkozunk vele termések falában is.
A szilárdító szövetek másik típusa a szklerenchima. Ennek elemeit erősen lignifikált sejtfal merevíti, ami a faanyag berakódása miatt olyan mértékű izolációt jelent a sejtek számára, ami azok pusztulását okozza. A szklerenchima két megjelenési formája a szklereida és a rost. A szklereidák változatos alakú sejtek, amelyeket többnyire elszórtan figyelhetünk meg a szövetekben. Csillag alakú aszteroszklereidák merevítik a vízi növények aerenchimáját, izodiametrikus brachiszklereidák (kősejtek) találhatók kis csoportokban elszórva a Pyrus [körte] fajok gyümölcshúsában, a csonthéjas termések endokarpiumát („csonttár”) szintén kősejtek építik fel, de megtalálhatók egyes szárak (pl. Hoya [viaszvirág]) kérgi régiójában is. Oszlop alakú makroszklereidák illetve csontra emlékeztető megjelenésű oszteoszklereidák különféle magvak maghéjában (pl. Phaseolus [veteménybab]) fordulnak elő.
A kis átmérőjű, szűk üregű, megnyúlt (gyakran több centiméter hosszúságú) szilárdító alapszöveti elemek a rostok. Két fő típusuk a xilem részét képező xiláris rostok és a háncselemekhez kapcsolódó extraxiláris rostok. Az előbbiek erősebben lignifikált, vastagabb falú rostok, a faelemekkel közös osztódószöveti sejtek leszármazottai. Lehetnek hosszú, megnyúlt, igen vastag falú libriform rostok vagy a tracheidákhoz hasonló, de azoknál vastagabb falú (lényegében a tracheida és a libriform rost közti átmenetet képviselő) rosttracheidák, amelyek a nyitvatermők legfontosabb szilárdító elemei. Valódi libriform rostokat csak a zárvatermő fajokban találunk (gyakran rosttracheidák mellett). Az extraxiláris rostok érdekessége, hogy faluk többnyire vékonyabb a xiláris rostokénál, hosszuk pedig jelentősen meghaladja azokét (a Boehmeria nivea [hócsalán] esetében akár az 55 cm-t is elérheti). A rostok szinte minden esetben szállítónyalábokhoz kapcsolódva, azok belsejében vagy körülöttük, nyitott vagy zárt nyalábhüvelyt képezve fordulnak elő a növényben. Az extraxiláris rostok kötegei gyakran mint „háncssapka” vagy „nyalábsapka” kísérik a nyalábokat a floem felőli oldalon.
Megjegyzendő, hogy az ipar nem az anatómia értelemben vett rostokat tekinti növényi rostoknak. Az esetek többségében az ipari rost kifejezéssel a szállítónyalábokat és az azokat körülvevő, rostokból álló nyalábhüvely összességét jelöljük. Hagyományos rostnövényeink közül a kender (Cannabis savtiva) háncsrostjait kötelek, zsinegek, durva vásznak készítéséhez használják, miután a farésztől áztatással majd tilolással megszabadítják. A len (Linum usitatissimum) háncsrostja tiszta cellulózból álló kéregrost, puha lenvászon készítéséhez használták. Ebbe a gazdasági kategóriába tartoznak azonban a gyapot (Gossypium) maghéjának szinte tiszta cellulózból álló epidermális szőrei is.
A kiválasztó alapszövet: a növények belső (endogén) kiválasztó képletei
Az epidermisz módosulatai és felszíni kiválasztó képletei mellett a növényi szervezet belsejében is találunk szekrécióra képes sejteket. Ezek az alapszöveti eredetű sejtek vagy elszórtan találhatók a növényi szervezetben (kiválasztó idioblasztok), vagy zsák- illetve hólyagszerű szekréciós üregeket, esetleg cső alakú szekréciós járatokat alkotnak.
A sejtszintű kiválasztás típusai
A szekréciós üregek és járatok abban különböznek az idioblasztoktól, hogy váladékuk a sejtek közötti térben halmozódik fel.
A váladéktartó üregrendszerek változatossága
Az összetettebb belső kiválasztó képletek közé tartoznak a legtöbb nyitvatermő fatestét és alapszöveteit átszövő, skizogén eredetű gyantajáratok, amelyek egyes zárvatermő családokban (pl. Anacardiaceae [szömörcefélék]) is előfordulhatnak. A citrusfélék (Rutaceae) termésfalában lizigén eredetű illóolajtartók fordulnak elő (11. ábra). E kiválasztó képletek váladéka csípős, keserű ízével a rovarrágástól védi a növényi szerveket.
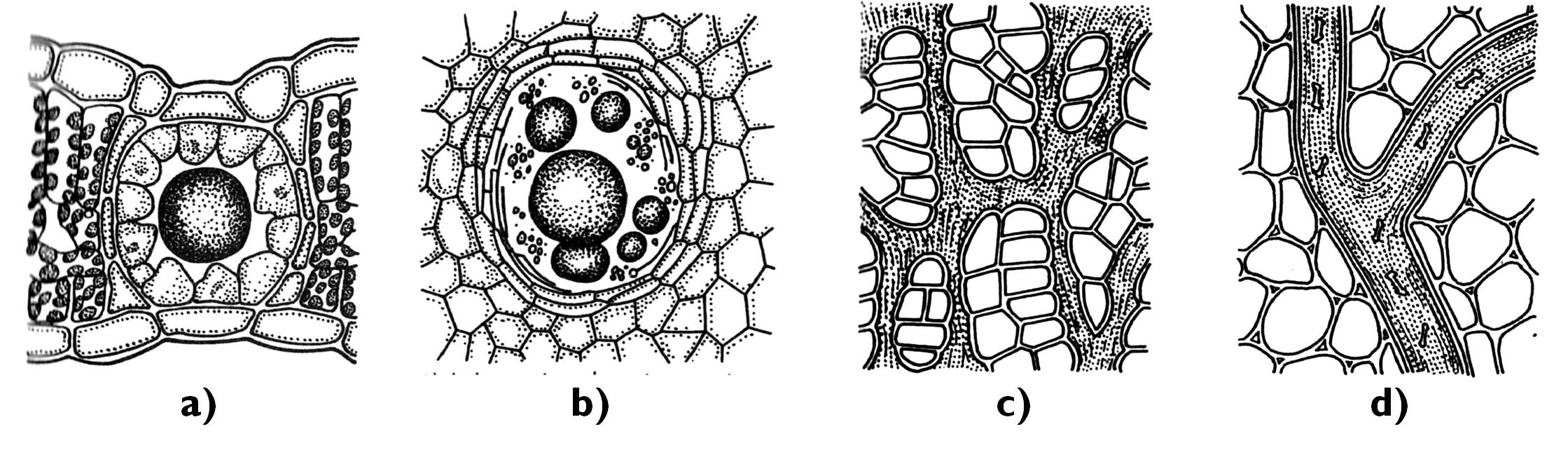
11. ábra. A belső kiválasztó képletek.
a) skizogén olajtartó (Hypericum); b) lizigén illóolajtartó (Citrus); c) tagolt tejcső (Scorzonera); d) tagolatlan tejcső (Euphorbia).
(Jacob és mts. nyomán)
A növényi szervezet számos szövetét behálózó belső kiválasztó képletek a tejcsövek (laticiferek). Eredetüket tekintve lehetnek egyszerű tejcsövek, ha egyetlen sejt kiterjedésével jönnek létre, vagy összetett tejcsövek, amennyiben több sejt összeolvadásával keletkeznek.
A tejcsövek sokfélesége és előfordulásuk
A tejcsövekben felhalmozódó anyagok összessége a tejnedv (latex), ami különféle anyagok (köztük lipidek) vizes oldatának illetve kolloidális emulziójának tekinthető. Lehet színtelen, vízszerű (Morus [eperfa], Humulus [komló], Nerium [leander]), tejszerű, fehér (Asclepias [selyemkóró], Euphorbia [kutyatej], Ficus [füge], Lactuca [saláta]) vagy színes (Cannabis [kender], Papaveraceae [mákfélék]). Alkotói különböző szénhidrátok (cukrok, keményítőszemcsék), szerves savak, sók, szterolok, zsírok, terpenoidok (pl. kaucsuk), alkaloidok, fehérjék (akár enzimek is – pl. a Carica papaya [papaja] esetében emésztőenzimek). Számos faj tejnedve fontos ipari alapanyag (pl. a Hevea brasiliensis [kaucsukfa] tejnedve, a kaucsuk) vagy gyógyászati hatóanyagforrás (pl. a Papaver somniferum [mák] tejnedvéből, az ópiumból ópiátokat nyernek, az Apocynaceae [meténgfélék] család tagjaiból a vinca-alkaloidokat). Számos dísznövényünk (pl. Nerium oleander, Vinca minor) mérgező tulajdonságát is magas alkaloid-tartalmú tejnedvének köszönheti.
Több szövetrendszer által kialakított szervfüggelékek – az emergenciák
A növényi szövetek és szervek közti átmenetet képviselő anatómiai képletek az emergenciák (szervfüggelékek). Ezek felépítésében a bőrszövet mellett egyéb szöveti elemek (alapszövet, néha szállító szövet) is részt vesznek, ezért egyik szövetrendszerhez sem sorolhatók be, de nem tekinthetők önálló szerveknek sem.
Emergencia például a Rosaceae [rózsafélék] számos tagjára (Rubus [szeder, málna], Rosa [rózsa]) jellemző tüske, vagy az Urtica [csalán] nemzetség csalánszőreinek alapszövetből álló, epidermisszel fedett alapi, párnaszerű része. Szintén ebbe a kategóriába sorolhatók a harmatfüvek (Drosera) rovarfogó tentákulumai. Ezek olyan nyeles mirigyek, melyek feji részét nyálkaanyagot és emésztőnedvet előállító kiválasztó epidermisz borítja, belső alapszöveti állományába pedig szállítóelemek (tracheidák, rostasejtek) ágyazódnak. Ez utóbbiak a kiválasztott anyagok előállításához szükséges vegyületek valamint a felszívott emésztési végtermékek szállítását végzik.
Ellenőrző feladatok
Ide jó lenne a feladatokat feltölteni
Irodalomjegyzék
Beck CB (2010): An Introduction to Plant Structure and Development: Plant Anatomy for the Twenty-First Century. 2. kiad. Cambridge University Press, Cambridge,
Biebl R, Germ H (1967): Praktikum der Pflanzenanatomie. 2. kiad., Springer-Verlag, Wien-New York.
Braune W, Leman A, Taubert H (1967): Pflanzenanatomisches Praktikum. VEB Gustav Fischer Verlag, Jena.
Cutler DF, Botha T, Stevenson DW(2008): Plant Anatomy: An Applied Approach. Wiley-Blackwell, Oxford.
Evert RF, Eichhorn SE (szerk.) (2006): Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body – Their Structure, Function, and Development, 3. kiad., Wiley-Interscience, New Jersey.
Gyurján I (1996): Növényszervezettan. Egyetemi jegyzet. ELTE Eötvös K., Bp.
Haberlandt G (1924): Physiologische Pflanzenanatomie. Verlag von Wilhelm Engelmann, Leipzig.
Haraszty Á (1998): Növényszervezettan és növényélettan. 6. kiad., Nemzeti Tankvk., Bp.
Jacob F, Jäger EI, Ohmann E (1985): Botanikai kompendium. Natura, Bp.
Kárpáti Z, Görgényi L, Terpó A (1968): Kertészeti növénytan I. Növényszervezettan. Mezőgazdasági Kiadó, Bp.
Mauseth JD (2008): Plant Anatomy. The Blackburn Press, New Jersey.
Sárkány S, Haraszty Á (1995): Növényszervezettan biológia-kémia, biológia ill. pszichológia-biológia szakos hallgatók részére. Egységes jegyzet. Nemzeti Tankvk., Bp.
Sárkány S, Szalai I (1966): Növénytani praktikum I. Növényszervezettani gyakorlatok. 3. kiad., Tankvk., Bp.
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028