Sejttan (Citológia)
Szerző: Erős-Honti Zsolt
A sejtfogalom és a sejtelmélet kialakulása, változása
Ma már egyértelműen elfogadott a tény, hogy valamennyi élőlény teste sejtekből áll. A 17. század közepéig azonban a kutatóknak nem volt egyértelmű tudomásuk arról, hogy minden szervezet kisebb építőelemekből állna. Az 1650-es években Jan Swammerdam holland természettudós fedezett fel ovális testecskéket a vérben, majd megállapította azt is, hogy a békák embriója apró gömböcskékből áll. Növényi sejteket elsőként Robert Hooke angol polihisztor írt le 1663-ban, fás szár paraszövetének vizsgálata során. Micrographia című munkájában a megfigyelt, méhek lépsejtjeire emlékeztető képződményeket „cellula” („kamrácska”, „üreg”) névvel illette: tőle származik tehát egyszersmind a sejtek megnevezése is. (Habár úgy vélte, hogy a „cellulák” vízszállításra szolgáló csövecskék, s később kiderült, hogy valójában nem is élő sejteket, hanem holt sejtek sejtfalait látta mikroszkópjában, a megnevezés azóta is megmaradt.)
Az első sejtleírások után több mint 150 évnek kellett eltelni ahhoz, hogy valamennyi élőlényre kiterjesszék a sejtes szerveződés elvét. 1838-ban osztotta meg megfigyeléseit egymással Mathias Jakob Schleiden német botanikus és Theodor Schwann német fiziológus: előbbi a növényi szervezet, míg utóbbi az állatok testével kapcsolatban állapította meg a sejtes szerveződés általános mivoltát. Eredményeiket 1839-ben Schwann könyvében tették közzé, ebben fogalmazták meg a klasszikus sejtelmélet pontjait is:
- Valamennyi élőlény anatómiai, élettani és szerveződési alapegysége a sejt;
- a sejt kettős természetű: egyszerre különálló, élő egység és a szervezet építőköve;
- a sejtek – a kristályokhoz hasonlóan – az élettelen anyagból szerveződnek.
Az utolsó pont alapján Schleiden és Schwann úgy vélték, hogy az élő sejt sejtmagja egy idő után kikristályosodik, a sejten kívüli térbe kerül, ahol kiterjed, majd belőle újra élő sejtek jönnek létre; az élettelen és élő fázisok tehát váltogatják egymást, s egyfajta folytonosságot képviselnek. Ez az ókor óta elfogadott spontán keletkezés („natura non facit saltus”, azaz „a természet nem végez ugrásokat”) elméletének továbbéléseként is felfogható. A klasszikus sejtelmélet 3. pontját később Robert Remak, Albert Kölliker és Rudolf Virchow sejtek szaporodási folyamataira irányuló kutatásai döntötték meg. Virchow mondta ki először azt a tényt, miszerint „minden élő sejt élő sejtek közvetlen leszármazottja” („omnis cellula e cellula”), ami azóta is elfogadott vélemény.
A modern sejtelmélet kidolgozása
A sejtek evolúciója: a prokarióta és az eukarióta sejt összevetése, az eukarióta sejt típusai
Az első élő sejtek megjelenésére vonatkozóan csak közvetett bizonyítékok állnak rendelkezésünkre. Ezek alapján kialakulásukat mintegy 3,8-4 milliárd évvel ezelőttre becsüljük. Habár az evolúcióbiológusok számos elméletet dolgoztak ki arra vonatkozóan, hogy milyen lépések során jöttek létre az ősóceánok vizében oldott szerves makromolekulákból az első élő sejtek, a molekulák egymásba alakulását jelentő prebiológiai evolúció és a biológiai evolúció közti határ átlépéséről mindmáig csak feltételezéseink vannak.
Az evolúció során elsőként megjelent élő sejtek felépítése még igen egyszerű volt. Napjainkban a különböző baktérium-csoportok sejtjeinek felépítésében figyelhetjük meg ezeket az ősi bélyegeket. A legfontosabb jellemzője ezeknek a sejteknek, hogy örökítőanyaguk még szabadon úszik a sejtplazmában, még nincsen valódi, sejtmaghártyával körülhatárolt sejtmagjuk. Ezen tulajdonságuk alapján ezeket az élőlényeket (és valamennyi ma élő baktérium-csoportot) prokariótáknak nevezzük, szemben a sejtmaghártyával határolt, valódi sejtmagvas eukariótákkal, amelyek kb. 2-3 milliárd éve alakultak ki. A sejtmag hiánya mellett számos egyéb különbséget is fel tudunk sorolni a két sejttípus között (ezeket az 1. táblázat összegzi): a prokarióta sejtben jellemzően nincs kiterjedt belső membránrendszer, nem tartalmaz nagyobb sejtorganellumokat, mérete néhány mikrométeres nagyságrendű az eukarióták 10-100 mikrométeres sejtjeivel szemben, s eltér a két sejttípus DNS-ének szerveződése is.
1. táblázat. A prokarióta és az eukarióta sejt összehasonlítása.
|
Prokarióta sejt |
Eukarióta sejt |
|
|
Megjelenés |
3,8 milliárd éve |
2,7 milliárd éve |
|
Méret |
~ 1 μm |
10-100 μm |
|
Sejtmag |
Nincs |
Van |
|
Örökítőanyag |
1 db, |
Mindig több, nyílt láncú (lineáris) DNS-molekula |
|
„Nem-hiszton” fehérjékhez kötött DNS |
Hisztonfehérjékhez kötött DNS |
|
|
1×106 ― 5×106 bázispár |
1,5×107 ― 5×109 bázispár |
|
|
Replikáció |
Egy ponton indul |
Több ponton indul |
|
Sejtciklus |
Nincs |
Van |
|
Osztódás |
Hasadás (amitózis) |
Mitózis, meiózis |
|
A kromoszómák mozgása a sejthártyához kötött |
A kromoszómák mozgása a magorsóhoz kötött |
|
|
Belső membránrendszer |
Kevés |
Kiterjedt (ER, Golgi stb.) |
|
Sejtváz |
Nincs (kezdetleges) |
Van |
|
Sejtorganellumok |
Kevés |
Mindig megvannak |
|
Mozgató sejtszervecskék |
Fehérjeszálak |
Valódi csilló, ostor |
|
Riboszómák mérete |
70S (30S + 50S) |
80S (40S + 60S) |
|
Transzkripció és transzláció |
Egy térben |
Térben elválasztva |
|
Sejtkapcsoló struktúrák |
Nincsenek |
Vannak |
|
Differenciálódási képesség |
Nincs (alacsony szintű) |
Van |
|
Citózis |
Nincs |
Van (lehet) |
|
Többsejtűség |
Ritkán (pl. kékbaktériumoknál) |
Többnyire többsejtűek |
|
Programozott sejthalál (apoptózis) |
Nincs (bizonytalan) |
Van |
Lynn Margulis elmélete az endoszimbiózisról
Annak ellenére, hogy alapvető felépítésükben megegyeznek, az eukarióta élőlények sejtjei is különböznek egymástól attól függően, hogy melyik eukarióta királyságba (Regnum) tartozó szervezetet alkotják (2. táblázat). A legszembetűnőbb különbség, hogy a növényi és gombasejtek állandó alakját és térfogatát biztosító sejtfal hiányzik az állati sejtek esetében. A sejtfal összetétele azonban eltérő a két királyságban: a növények sejtfalában található legfontosabb poliszacharid a cellulóz, míg a legtöbb gombasejt esetében ez valamilyen – az állatvilágban is előforduló – kitinhez hasonló vegyület. Az állati sejtek általában rendelkeznek a helyváltoztatás képességével: állábakkal, csillókkal vagy ostorok segítségével mozoghatnak. A gombák körében már ritkábban, de még mindig több esetben találkozunk ostoros sejtalakokkal (zoospórákkal, ivarsejtekkel vagy az ostorosgombák vegetatív sejtjeivel), viszont a növények körében csak egyes hímivarsejtek képesek aktív mozgásra. (A zárvatermők körében már ezt sem figyelhetjük meg.) Minden eukarióta sejtben találhatók kisebb-nagyobb méretű, folyadékkal telt ún. sejtüregek (vezikulumok), amelyek a sejtalkotók közti anyagáramlást biztosítják a makromolekulák számára a különféle „csomagolt”, azaz membrán közvetítette transzport-folyamatok során ill. a citózisokban. A növények és a gombák esetében ezek származékaként nagyméretű, elsősorban térkitöltésre és raktározásra szolgáló vakuólumok is megtalálhatók (amelyek folyadéktartalmából akár kristályzárványok is kiválhatnak). A lebontási folyamatok és az anyagok újrahasznosítását az állati sejtben az ún. lizoszomális rendszer tagjai biztosítják. (Újabban egyes növényi és gombasejtekben is bizonyították már hasonló szerepet betöltő ún. lítikus vakuólumok előfordulását.)
Alapvetően különbözik egymástól a három többsejtű eukarióta királyság képviselőinek anyagcseréje. A növények – fényenergiát hasznosítva – szervetlen anyagokból is képesek előállítani saját szerves anyagaikat: autotróf szervezetek. Ezzel szemben a gombák és az állatok sejtjei csak szerves vegyületekből képesek szerves anyagot (átépítéssel) előállítani, azaz heterotrófok (+info). Az autotróf anyagcsere szolgálatában állnak a csak növényi sejtekre jellemző színtestek, amelyek a többi eukarióta sejttípusból hiányoznak.
Csak gombákra jellemző sejtalkotók
2. táblázat. A növények, gombák és állatok sejtjeinek összehasonlítása.
|
Növényi sejt |
Gombasejt |
Állati sejt |
|
|
Szén-anyagcsere |
autotróf |
heterotróf |
heterotróf |
|
Sejtfal |
van, cellulóz-alapú |
van, kitin-alapú |
nincs |
|
Mozgató sejtszervecskék |
csilló, ostor |
ostor lehet |
csilló, ostor, álláb |
|
Jellemző s ejtüregtípus |
vakuólum |
vakuólum |
lizoszomális rendszer |
|
Speciális sejtalkotók |
színtestek |
csúcsi test, lomaszóma |
sejtközpont |
Animáció az eukarióta sejt felépítéséről . a mellékelt vonalrajz megfelelő sejtalkotóira ráállva megjelenik a sejtalkotó neve, majd funkciója. (Jacob és mts. nyomán)
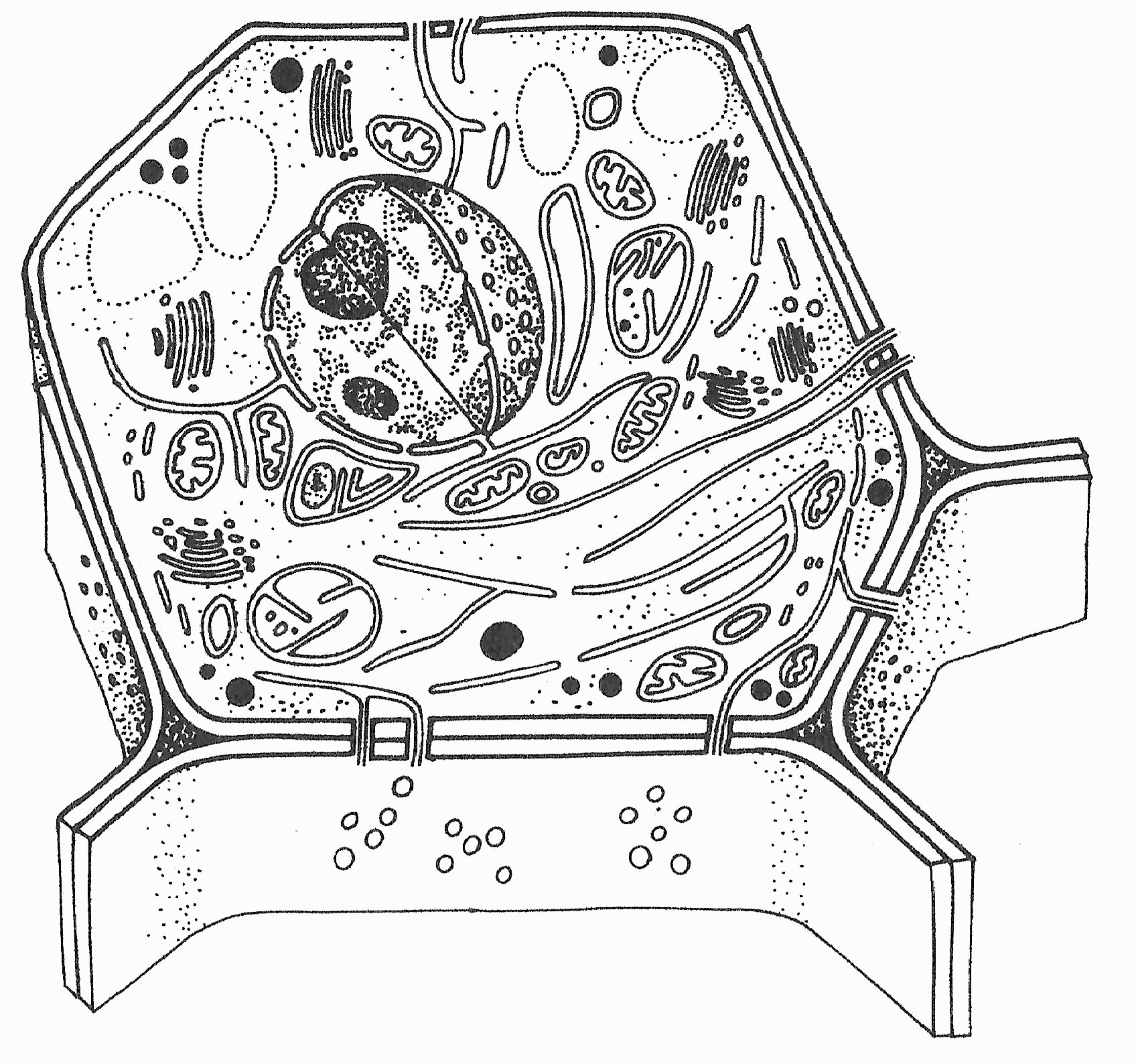
A sejtplazma és a sejtváz
Valamennyi élő sejt alapállományát több fázisú vizes oldat, a sejtplazma (citoplazma) képezi. Benne különböző ionok, köztes anyagcsere-termékek (cukrok, aminosavak, karbonsavak) és makromolekulák (enzimfehérjék, nukleinsavak) találhatók oldott állapotban. A felsorolt vegyületek különböző mérete miatt a sejtplazma egyszerre tekinthető valódi és kolloid oldatnak, sőt a sejtalkotók 1000 nanométert meghaladó mérete miatt durva diszperz rendszernek is. A sejtplazma egyes részein (többnyire a sejthártya alatti részen) a fehérjék hidrátburkai összekapcsolódnak, kocsonyás állapotúvá (géllé) alakítva a plazmát – ezt nevezzük citogélnek (megkülönböztetve a hígan folyó citoszoltól).
A sejtplazmában szabadon oldva találjuk egyes meghatározó anyagcsere-folyamatok enzimeit. Itt zajlik a monoszacharidok lebontásának (biológiai oxidáció) első szakasza, a glikolízis és az anaerob erjedés (fermentáció) is. A makromolekulák anyagcseréjének lépései közül a legfontosabb itt zajló folyamat a transzláció (fehérjeszintézis). A folyamat a fehérjékből és RNS-molekulákból (rRNS) felépülő makromolekuláris komplexek, a riboszómák felületén megy végbe. A riboszómák két részből, egy kis és egy nagy alegységből állnak, s feladatuk a hírvivő RNS (mRNS) információtartalmának (azaz nukleotid-sorrendjének) a fehérjék (pontosabban a polipeptidláncok) aminosav-sorrendjévé „fordítása”. A riboszómák kis alegységén találjuk az mRNS megkötéséért felelős felületet, míg a nagy alegység rRNS-molekulái katalizálják (mint katalitikus RNS-ek, azaz ribozimek) az aminosavak közti peptidkötések kialakulását. A riboszómákhoz az egyes aminosavakat a szállító funkciójú transzfer-RNS-ek (tRNS) szállítják.
A sejtplazmában oldott fehérjék között nemcsak enzimeket, hanem a sejt belső szerkezetét meghatározó fehérjéket is találunk. Ezek összessége a sejtváz (citoszkeleton) (1. ábra). A sejtváz határozza meg a sejtfallal nem rendelkező sejtek alakját, belső rendezettséget ad a sejtnek, hozzá köthetők a sejten belüli mozgások és a sejt mozgási jelenségei is. A sejtvázfehérjék gömbölyded alakú (globuláris) fehérjék, amelyek hosszú fonalakká kapcsolódnak össze. Jellegzetes sejtvázfehérje a merev, csőszerű kötegeket (mikrotubulusokat) létrehozó tubulin, vagy az elsősorban izomsejtek kapcsán ismert, de minden sejtben előforduló aktin is. A sejtvázfehérjékhez mind a két rendszer esetében a fehérjeszálakon (ATP-bontás terhére) elmozdulni képes ún. motorfehérjék kapcsolódnak.
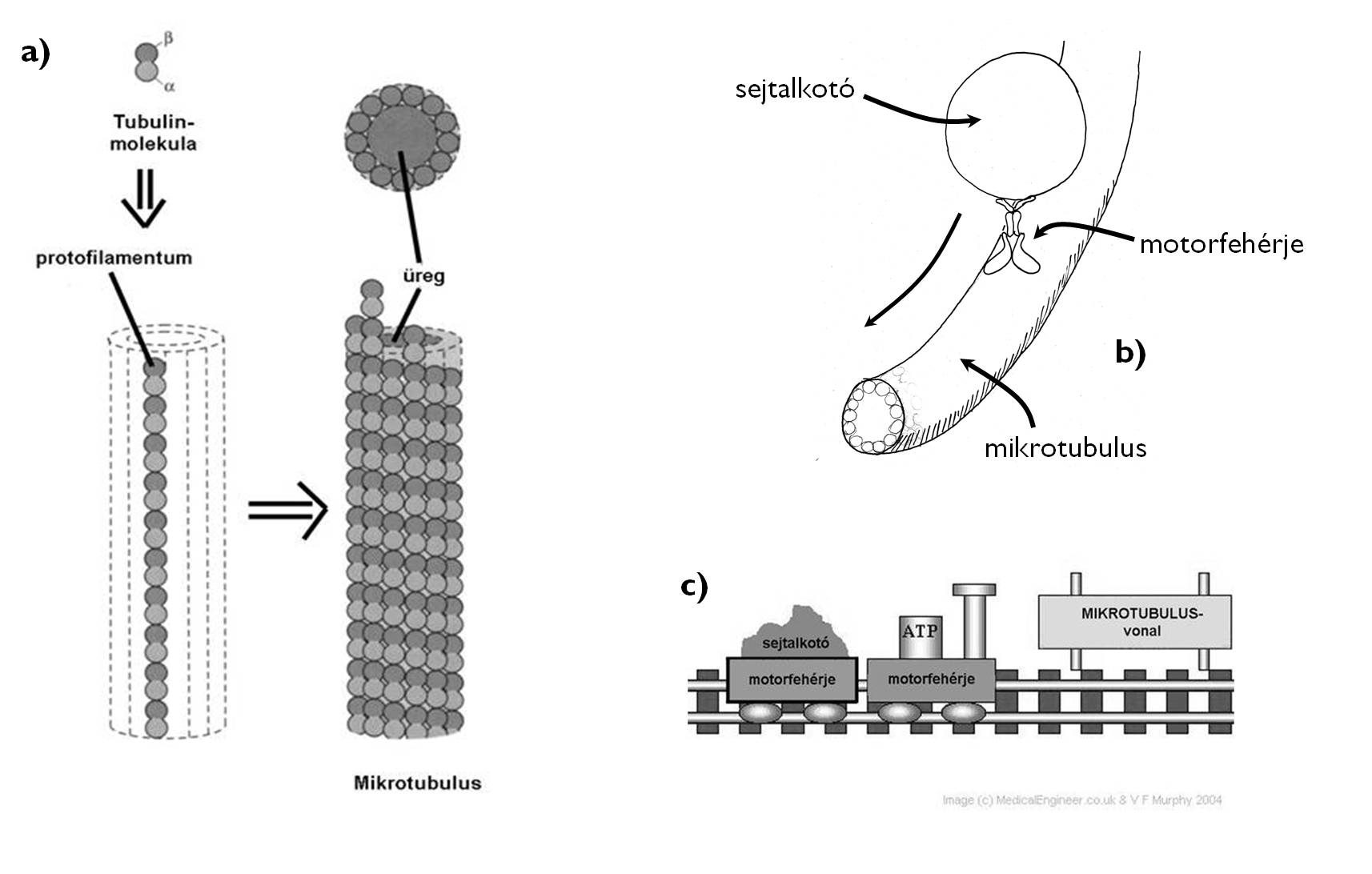
1. ábra. A mikrotubulusok felépítése (a) és a motorfehérjék működésének alapja (b, c).
(a, c: internetes források nyomán; a - http://www.studyblue.com, c - http://www.medicalengineer.co.uk)
A sejtváz típusai: a mikrotubuláris és az aktinváz
A sejten belüli mozgások irányítása mellett a citoszkeleton felelős a helyváltoztatásra képes sejtek mozgásaiért is. A csillók és ostorok egyaránt mikrotubulusokkal merevített plazmanyúlványok, amelyeket teljes felületükön a sejthártya borít be (2. ábra).
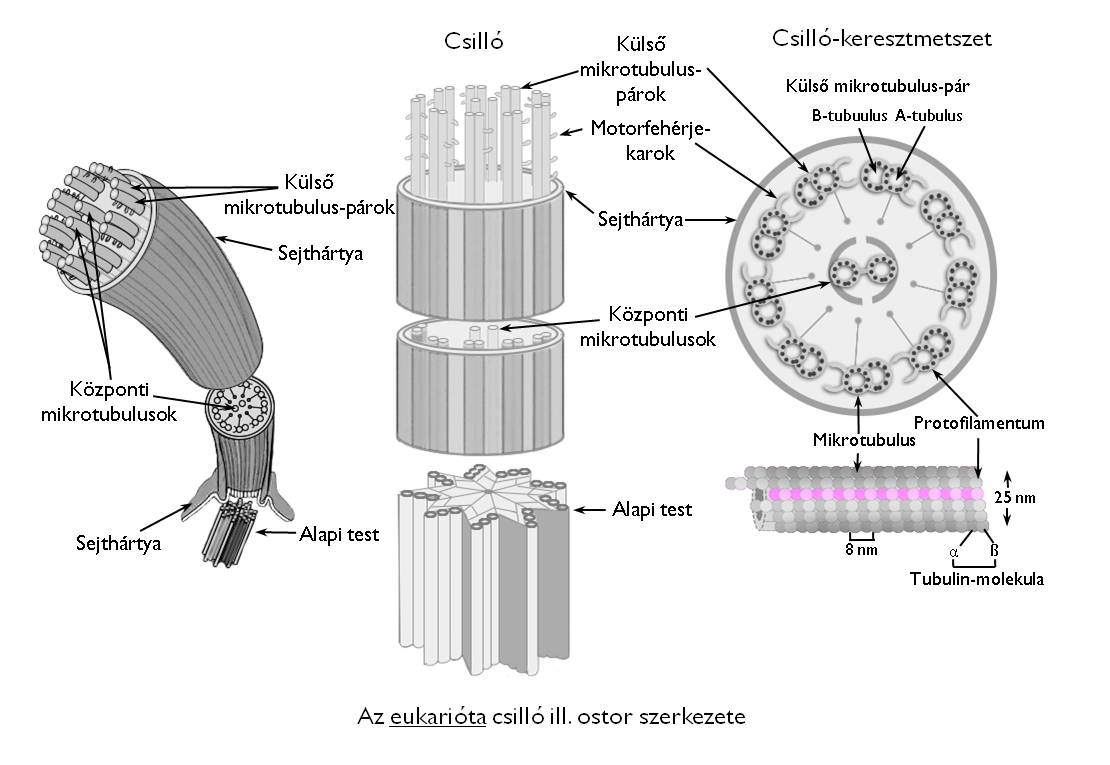
2. ábra. Az eukarióta csilló (ostor) felépítése. (Internetes forrás nyomán: http://scienceblogs.com)
A csilló és az ostor felépítése, működése
A sejt membránrendszerei
Az eukarióta sejtek belsejében szinte minden esetben kiterjedt belső hártyarendszert találunk. Ennek egyik feladata, hogy különböző összetételű térrészekre (kompartmentekre) tagolja a sejt belső terét, így elkülöníthetővé válnak az egymást zavaró anyagcsere-folyamatok, illetve elválaszthatók egymástól olyan anyagcsere-termékek, amelyek a sejt számára hátrányos reakcióba léphetnének egymással. (Ezt a jelenséget nevezzük kompartmentalizációnak.) A belső membránrendszerek kialakulásának másik evolúciós előnye, hogy a hozzájuk kötődő enzimek révén kiterjedt felületet biztosítanak egyes biokémiai reakciók számára.
A biológiai membránok felépítése: a folyékony-mozaik membránmodell
A sejthártya és a transzportfolyamatok
Valamennyi élő sejtet egy lipid kettősréteg alapú membrán, a sejthártya (plazmalemma) határol el környezetétől. A sejthártya feladata kettős: elhatárolja a sejt protoplazmáját a külvilágtól, emellett biztosítja a sejt és a külvilág kapcsolatát. A kapcsolattartás részeként a sejthártya speciális molekulái biztosítják a sejtek egyedi felismerhetőségét. Emellett a külvilágból jövő anyagok és a sejt által termelt vegyületek is a sejthártyán keresztül mozognak a belső és a külső tér között a különféle transzportmechanizmusok segítségével (3. ábra). Csak a növényi sejtre jellemző speciális funkcióként a sejthártyába ágyazódva találjuk a sejtfal cellulózkomponensének előállítását végző enzimeket is.
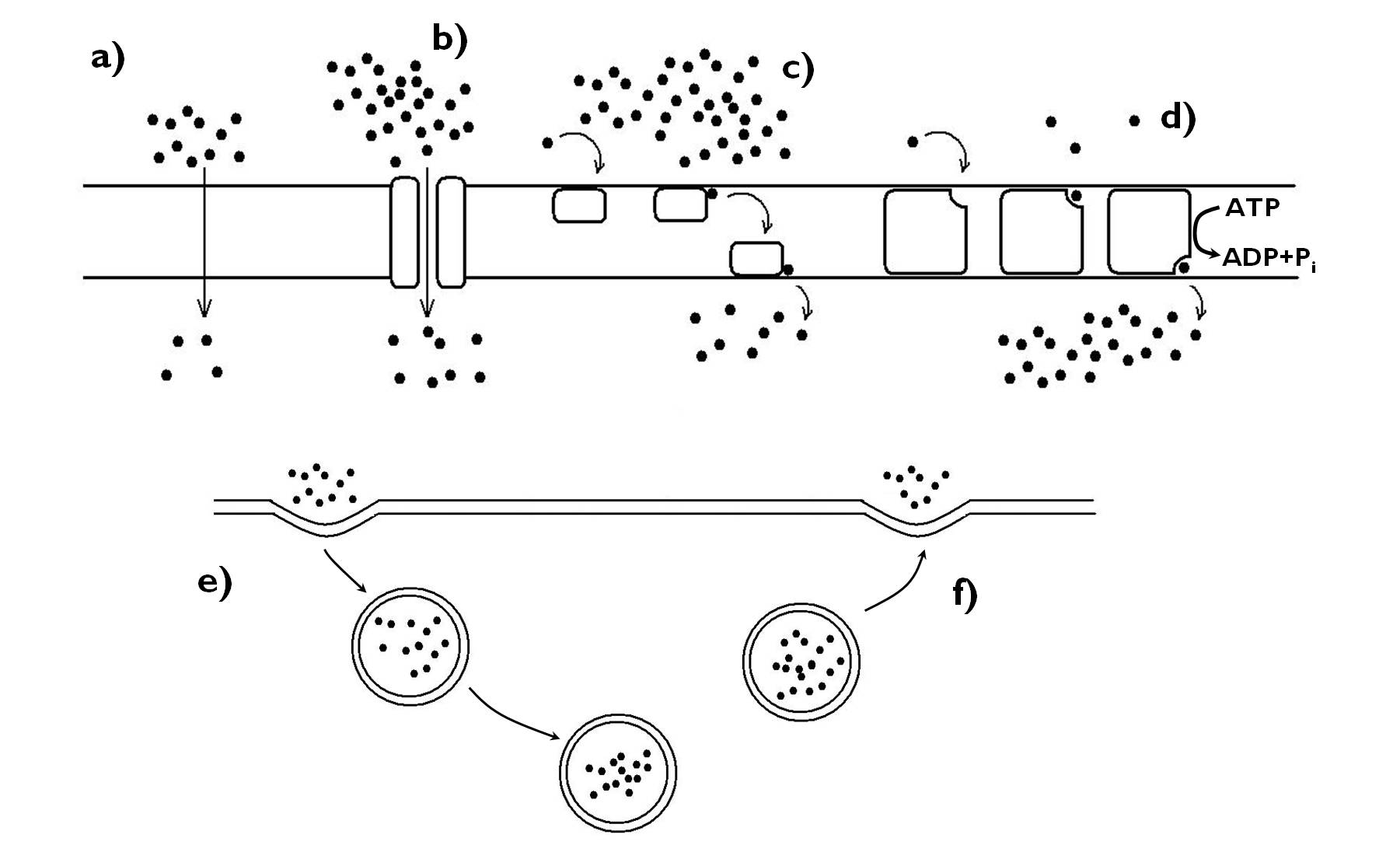
3. ábra. A membránokon keresztül folyó transzportok.
a) szabad transzport (diffúzió), b-d) közvetített transzport (b: csatornafehérje, c: hordozófehérje, d: pumpafehérje), e)-f) citózis (e: endocitózis, f: exocitózis).
A sejt szekréciós apparátusa (ER, Golgi)
Az eukarióta sejt belsejében előforduló membrán-sejtalkotók egyik fontos feladata, hogy egymással együttműködve előállítsák a sejt leadásra szánt anyagait. Ezt a folyamatot nevezzük elválasztásnak vagy szekréciónak, a folyamatban résztvevő sejtalkotó-együttest pedig a sejt szekréciós apparátusának (4. ábra). A szekréciós apparátus tagjai az endoplazmatikus hálózat, a Golgi-készülék, a sejthártya és a felsorolt organellumok közti kapcsolatot biztosító membrán-vezikulumok.
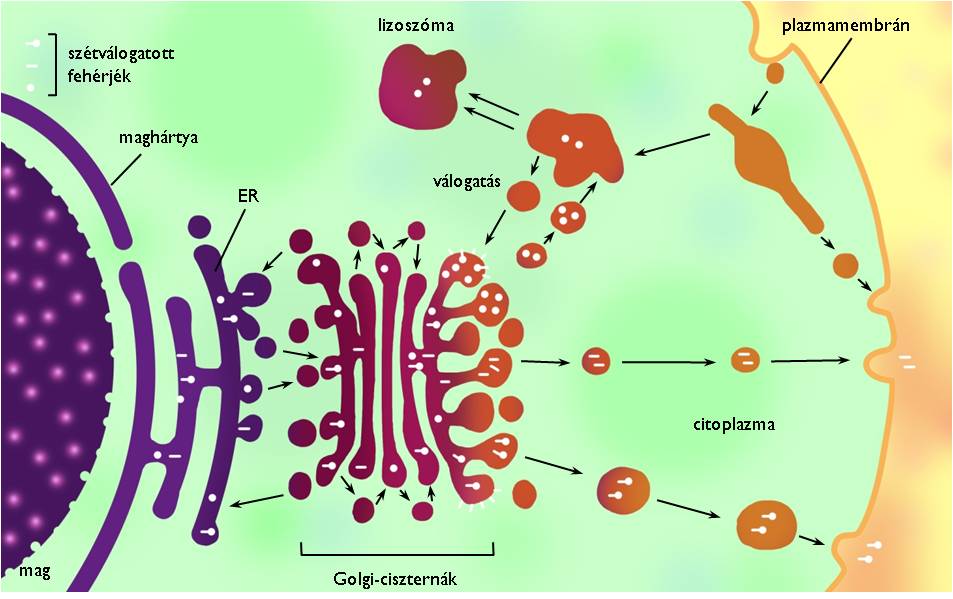
4. ábra. Az eukarióta sejt szekréciós és lizoszómális apparátusa. (Kovács nyomán)
Az endoplazmatikus hálózat (endoplazmatikus retikulum, ER) egymással folytonos kapcsolatban álló, kiterjedt membránzsákok (ún. ciszternák) rendszere a sejtplazmában. Két típusa létezik, a durva felszínű ER (DER) és a sima felszínű ER (SER). A DER felületéhez riboszómák kapcsolódnak, ezért elektronmikroszkópos felvételeken szemcsézettnek, rücskösnek látszik. A riboszómák – a sejtplazma szabad riboszómáihoz hasonlóan – fehérjeszintézist végeznek, a DER felületén azonban a sejtből leadásra szánt fehérjék jönnek létre. A folyamat során a keletkező fehérjék bekerülnek a DER üregrendszerébe (azaz a DER membránja elhatárolja őket a sejtplazmától). A DER folytonos kapcsolatban áll a sejt örökítőanyagát körülvevő sejtmaghártyával is, ami egyik bizonyítéka annak az elméletnek, miszerint az evolúció során a magmembrán a DER kiterjedésével jött létre. A SER elsősorban a lipidszintézis (pl. szteroid-, foszfolipid-előállítás) és a méregtelenítés helye, de – amennyiben szükség van rá a sejten belül – részt vesz a kalciumionok raktározásában is.
Az ER-ben keletkezett vegyületek az ER-zsákok kitüremkedésével kialakuló membránhólyagok (vezikulumok) belsejébe kerülve jutnak el a szekréciós apparátus következő tagjához, a Golgi-készülékhez (Golgi-apparátus). A Golgi egymás fölött elhelyezkedő, független membránzsákok és az azok közti kapcsolatot megteremtő vezikulumok együttese. (Ezeknek a membránzsákoknak a mérete jóval kisebb az ER ciszternáinál.) A sejtalkotó feladata a leadásra szánt fehérjék módosítása (elsősorban szénhidrát-oldalláncokkal történő ellátása), a külvilágba kerülő poliszacharidok szintézise (a cellulóz kivételével!) valamint a szekretálandó anyagok osztályozása is (+info). A leadásra szánt anyagok szekréciós vezikulumokba csomagolva hagyják el a Golgi-készüléket. Ezek végül a sejthártyához vándorolnak (a sejtvázfehérjéken mozgó motorfehérjék segítségével), ahol végül exocitózissal ürítik a külvilágba tartalmukat.
A vakuólum és a kristályzárványok
A növényi (és gomba-)sejtek jellegzetes membránnal határolt sejtalkotói a folyadékkal telt vakuólumok (sejtüregek). Ennek a sejtalkotónak az elsődleges feladata olyan, folyadék természetű vegyületeknek a raktározása és elkülönítése, amelyek jelenlétükkel zavarnák a sejtplazmában zajló anyagcsere-folyamatok enzimeit. Ilyen vegyületek például a különféle karbonsavak (almasav, citromsav – gyakran sóik formájában). A vakuólum tartalma a sejtnedv.
A differenciálódott sejtek az esetek többségében kevesebb (ált. egyetlen), nagyméretű vakuólumot tartalmaznak. Ennek a hátterében az áll, hogy a növényi sejtek növekedése intenzív vízfelvétellel valósul meg (a belső, ún. turgornyomás feszítő ereje tágítja a differenciálatlan sejtek még képlékeny sejtfalát). A felvett víz azonban annyira felhígítaná a sejtplazmát, ami már akadályozná a biokémiai folyamatok végbemenetelét, így a felvett víztöbbletet a sejt egy membránnal (az ún. tonoplaszttal) határolja el a sejtplazmától. (Tehát maga a víz is a vakuólumban raktározott folyadéknak tekinthető.) Az is előfordulhat, hogy a vakuólumban nem folyadék, hanem szilárd anyag, pl. fehérje-kristályok raktározódnak – ez az ún. aleuronszemcsék képződésének egyik lehetősége. (A vakuólum mellett raktározott fehérjék a DER-en is keletkeznek.)
A vakuólumban levő sejtnedvben találjuk a lilás-vöröses sejtek színét adó, pH-változástól függően színváltozásra képes színanyagokat, az antociánokat is (pl. a lilahagyma húsos allevelének epidermisz-sejtjeiben vagy a lilakáposzta-levelek bőrszövetében). Mindemellett a benne levő bontóenzimek révén a vakuólum végzi a sejten belül zajló hidrolitikus (emésztési) folyamatokat is (azaz ez a sejtalkotó látja el a növényi sejtekben a lizoszomális rendszer feladatát).
Mivel a növények nem végeznek az állati szervezetben zajló kiválasztáshoz hasonló folyamatokat, sejtszinten különítik el a felvett méreganyagokat, továbbá a felesleges ill. sokszor káros anyagcsere-termékeiket is. Ennek az elkülönítésnek egyik lehetősége, ha a káros vegyületeket (szerves savakat) a sejt oldhatatlan csapadékká alakítja. Ennek a színtere is a sejtnedv-vakuólum. Ebben az esetben azonban Ca2+-ionok kerülnek a vakuólumba, és az ott levő szerves savakkal oldhatatlan sókat (pl. CaCO3 /mészkő/ vagy Ca(COO)2 /kalcium-oxalát/ kristályokat) hoznak létre. Az így kialakuló kristályokat hívjuk zárványnak (5. ábra). A szerves savak azonban maguk is szerepet játszhatnak további méreganyagok semlegesítésében: szerves sókat képezhetnek pl. nehézfém-ionokkal - malát, citrát, oxalát formájában.
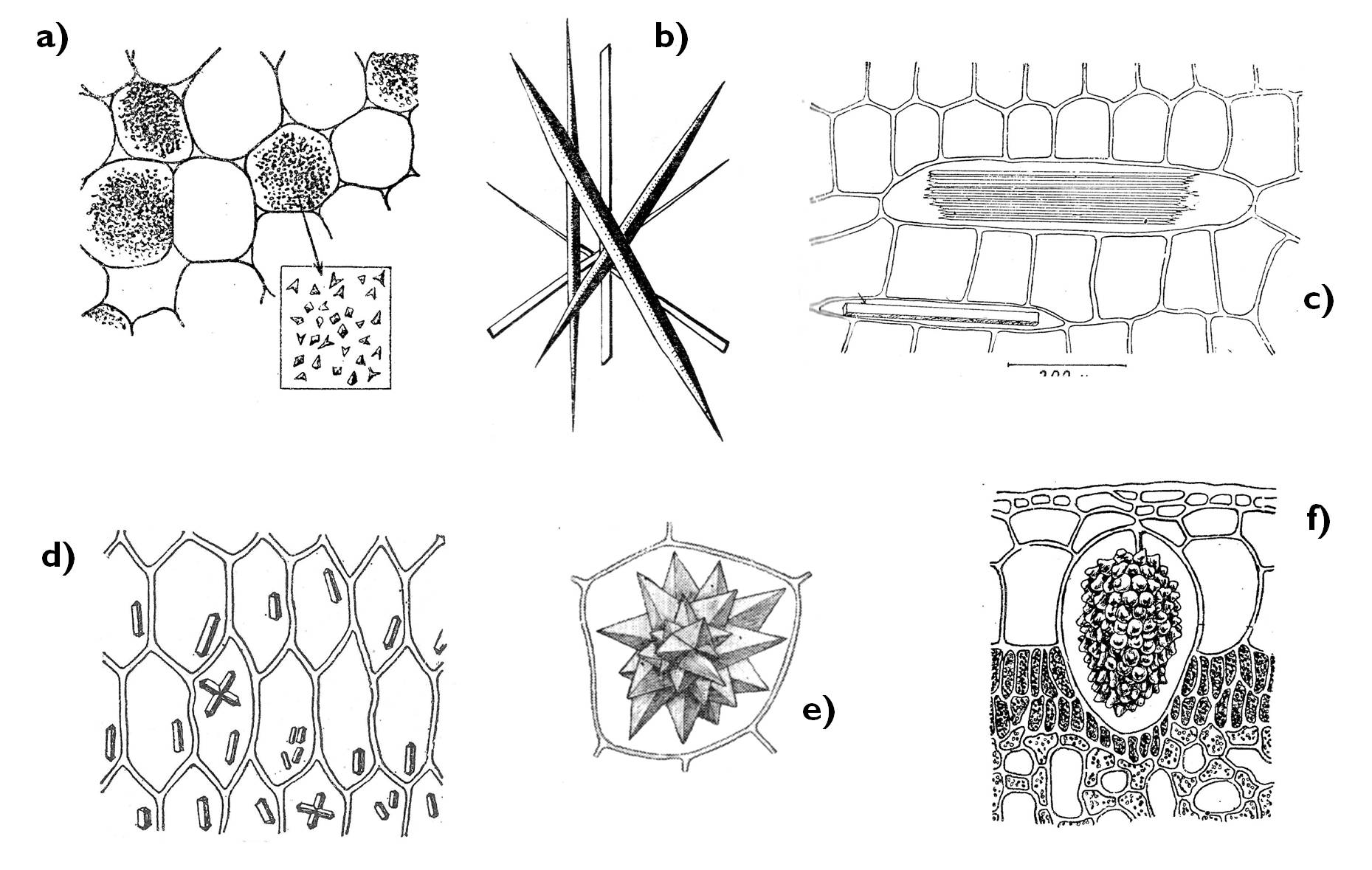
5. ábra. A kristályzárványok típusai.
a) kristályhomok, b) tűkristályok (sztiloid), c) tűkristály-köteg (rafid), d) oszlopkristályok, e) rozetta-kristály (drúza), f) fürtkristály (cisztolit). (Sárkány és Szalai nyomán)
A sejtfal
A sejthártyán kívül, a sejtek többsége esetében egy többé-kevésbé szilárd, merev sejtalkotó, a sejtfal helyezkedik el. Alapvetően hasonló funkciójú és szerveződésű (de rendszertani csoportonként eltérő molekulákból felépülő) sejtfal veszi körül a baktériumok, algák, gombák és növények sejtjeit. A növényvilágban igen ritkák az olyan sejtek, amelyeknek egyáltalán nincsen sejtfaluk. Ezeket a sejteket vagy sejtfallal rendelkező sejtek övezik, vagy pedig többnyire igen rövid életűek (pl. az ivarsejtek). Kivételt képeznek még a molekuláris-nemesítési vizsgálatokban gyakori modellszervezetek olyan növényi sejtjei, amelyeket enzimatikus kezelés révén fosztanak meg ideiglenesen sejtfaluktól – ezek az ún. protoplasztok.
A sejtfal elsősorban védelmi szerepet betöltő sejtalkotó. Védelmet nyújt a külvilágból érkező mechanikai hatásokkal és egyes kémiai vegyületekkel szemben, de a paraziták vagy a növényevő rovarok távoltartását is segítheti. Fontos szerepe, hogy a nagy mennyiségű vízfelvétellel növekvő növényi sejtek esetében védelmet nyújtson a sejtplazma felől jövő intenzív nyomással (ún. turgornyomással) szemben. Viszonylagos szilárdságának köszönhetően a sejtfal megszabja a növényi sejt alakját és méretét (térfogatát) is. Megjegyzendő mindazonáltal, hogy a sejtfal is képlékeny sejtalkotó, ami reagálni képes a környezet megváltozására vagy a sejt felől érkező hatásokra: a benne található enzimfehérjéknek köszönhetően folyamatos változásra képes. Éppen ezért nem tekinthetjük a sejt élettelen alkotórészének.
Egyes raktározó funkciójú növényi szervek esetében (pl. bizonyos fajok magjának táplálószövetében) a vastag sejtfal poliszacharid-komponense később tartalék-tápanyagként is mobilizálható a sejtfal visszabontása révén. A sejtfal tehát akár raktározásra is szolgálhat.
A növényi sejtfal kialakulása
A sejtek az osztódás (mitózis illetve meiózis) végszakaszában, a citokinézis során kettéfűződnek. A sejthatár létrejöttével párhuzamosan kerül sor a régi sejtfalakra merőlegesen az új sejtfal-szakasz kialakulására. (Az utódsejtek fala tehát nagyrészt megegyezik a kiindulási sejt sejtfalával, és csak az utódsejtek határfelületén alakul ki teljesen új sejtfal.)
Az elsődleges sejtfal kialakulásának folyamata
Nagy nagyítású mikroszkóppal vizsgálva a sejtfal rétegzett szerkezetet mutat. A szomszédos sejteket egy közöttük elhelyezkedő, elsősorban pektinből felépülő réteg, a középlemez kapcsolja egymáshoz. A középlemezzel szomszédos sejtfalréteg az osztódást követően elsőként kialakuló, ún. elsődleges sejtfal. Az elsődleges sejtfal vékony, képlékeny, még tágulékony; a differenciálódó (pl. osztódószöveti) sejteknek csak elsődleges sejtfaluk van. Később, a sejtek differenciálódásával az elsődleges sejtfalra további rétegek rakódnak rá: ezek a másodlagos sejtfal rétegei. A többnyire háromrétegű másodlagos sejtfal már jóval vastagabb, merevebb, további tágulásra már nem képes.
A végszakaszban kialakuló sejtlemez nem teljesen folytonos: néhány helyen pórusok törik át, amelyen az ER ciszternái húzódnak keresztül. A későbbi sejtfalrétegek kialakulásával is fennmaradnak ezek a pórusok, amelyek mentén a szomszédos sejtek sejtplazmája folytonos kapcsolatban áll egymással. Ezek a plazmahidak az ún. plazmodezmák.
Animáció a sejtfal kialakulásáról . az animációban a Golgi-készülékből származó, fragmoplaszt mentén mozgó vezikulumok összeolvadása, a sejtlemez kialakulása látható. Ezt követően a kialakuló középlemezre rárakódik az elsődleges sejtfal, majd erre a másodlagos sejtfal rétegei. (Az utódsejtek között plazmahidak maradnak – a plazmodezmák.)
A növényi sejtfal összetétele
A sejtfal szerkezete valamennyi e sejtalkotóval rendelkező rendszertani csoportban ugyanazt az általános felépítést követi: egy hidrogénkötésekkel stabilizált váz-poliszacharid (baktériumok esetében murein, gombák sejtjeiben kitin jellegű vegyületek, növényeknél cellulóz) ágyazódik bele a hozzá szintén hidrogén-hidak révén kapcsolódó makromolekulákból (további poliszacharidokból, fehérjékből és egyéb, speciális anyagokból) álló mátrixba. A sejtfal a szerkezeti vegyületek mellett enzimfehérjéket is tartalmaz, amelyek a sejtfalanyagok átépüléséért, a sejtfal megnyúlásáért, változásaiért és védekezésben történő közreműködéséért felelősek.
A növényi sejtfal vázát a több ezer ß-D-glükóz egységből felépülő nyílt láncú, lineáris cellulóz alkotja. Az egymás mellett, párhuzamosan futó cellulóz-láncok között erős hidrogén-hidak jönnek létre, ami egy kristályszerű szerkezetbe rendezi a fonalakat. (A rendezett szakaszokat, az ún. micellákat kevesebb hidrogénkötéssel merevített, rendezetlen („parakristályos”) cellulóz-szakaszok kötik össze.)
A sejtfalat alkotó cellulóz szintézise
A sejtfal alapállománya az ún. mátrix. Ezt egyrészt különféle poliszacharidok alkotják. Ezek közé tartoznak a hemicellulózok is, amelyek változatos monoszacharid-egységekből (glükóz, arabinóz, xilóz, mannóz stb.) álló, rövidebb láncú szénhidrátok. Hidrogénkötésekkel kapcsolódnak a cellulóz-mikrofibrillumokhoz és mintegy beburkolják azokat. A teljes sejtfal-mátrixot átitatják az ún. pektinek, amelyek szintén a poliszacharidok közé tartoznak. Elsődleges feladatuk, hogy egymáshoz tapasszák a sejtfal egyéb anyagait (ez a magyarázata annak is, hogy a szomszédos sejtfalakat összekapcsoló középlemez is pektinekből áll). A hemicellulózok és a pektinek a Golgi-készülékben keletkező, a sejtfalba exocitózissal ürülő poliszacharidok.
A sejtfal további poliszacharidjai: hemicellulózok és pektinek
A sejtfalban a poliszacharidok mellett poliszacharid-oldalláncokat tartalmazó fehérjéket (glikoproteideket) is találunk. Ezek egy része a sejtfal megnyúlásáért felelő enzimfehérje (pl. expanzin, hidrolázok, peroxidázok, transzferázok). Másik részük a sejtfal szerkezetének felépítésében részt vevő, a szénhidrátokhoz hidrogénkötések révén kapcsolódó struktúrfehérje (pl. az extenzinek).
Az eddig felsorolt komponensek (cellulóz, hemicellulózok, pektinek, fehérjék) mind az elsődleges, mind pedig a másodlagos sejtfalban előfordulnak. A másodlagos sejtfal jelentősebb vastagsága egyrészt annak köszönhető, hogy nagyobb mennyiségben tartalmazza az elsődleges sejtfal anyagait. Benne csak kisebb változások érintik ezen vegyületeket: megváltozik az egyes összetevők aránya, illetve a másodlagos sejtfalban a cellulózszálak lefutása párhuzamos egymással (szemben az elsődleges sejtfalban megfigyelhető rendezetlen lefutással).
Az elsődleges sejtfalanyagok mellett azonban a másodlagos sejtfal további, csak itt előforduló vegyületeket is tartalmaz. A legfontosabb ezek közül a lignin (faanyag). A lignin aromás vegyületek (kumarinsav, sziringol, ferulasav, galluszsav stb.) térhálós polimerje, melynek bizonyos összetevői rendszertani bélyegként is szolgálhatnak. Ez a térháló – a vasbetonban levő acélhálóhoz hasonlóan – átszövi a fásodott sejtfalat, fokozott szilárdságot, merevséget adva neki. (A lignifikálódott falú sejteket mondjuk „elfásodott”-nak.) A ligninberakódás elszigeteli a sejtet a környezetétől, olyannyira, hogy már tápanyagokat sem képes felvenni: az elfásodás tehát idővel a sejt halálát jelenti – pl. a szállítószövet farészében vagy a szklerenchima-rostok esetében.
A másodlagos sejtfal további speciális anyagai
A plazmodezmák
A növényi szervezet víztartalma alapvetően két különböző térrészben, az apoplasztban és a szimplasztban helyezkedik el. Az apoplaszt az élő sejtek protoplazmáján (azaz sejthártyáin) kívül elhelyezkedő „élettelen víztereket” jelöli: a sejtfal üregrendszerét és a sejtek közötti járatrendszert. A szimplaszt ezzel szemben az élő sejtek sejtplazmáiból álló „élő vízterek” összességének megnevezése. A szomszédos sejtek sejtfalainak elhelyezkedéséből fakadóan az apoplaszt vízterei magától értetődően többé-kevésbé összefüggő rendszert alkotnak a növényi testben. (Kivételt jelent ez alól a gyökér kérgi és sztélei régióinak apoplasztja, melyek között az endodermisz szuberinizált sejtfalai átjárhatatlan határt képeznek. – Részletesebben lásd a gyökér szövettanával foglalkozó fejezetben!)
Habár első benyomásra nem feltételeznénk az egyes sejtek szimplasztjai között átjárható kapcsolatot a növényi testben, mégis elmondható, hogy – az apoplaszthoz hasonlóan – a növényi test közel egységes szimplasztikus rendszert is alkot. A szomszédos sejtek szimplasztjait a sejtfal rétegein átnyúló citoplazma-hidak, a plazmodezmák kötik össze.
A plazmodezmák finomszerkezete és működése
A sejtfalvastagodás típusai
A differenciálódó sejtek nagy része esetében a másodlagos sejtfal rétegei az elsődleges sejtfal belső felületére rakódnak. Ebben az esetben a vastagodó sejt maga állítja elő a másodlagos sejtfal anyagait; a differenciálódással a sejtüreg mérete lecsökken. Ez az ún. centripetális sejtfalvastagodás (pl. a faelemek esetében). Ritkábban az is előfordul, hogy a sejtet körülvevő egyéb sejtek szintetizálják a másodlagos sejtfalanyagokat – pl. a pollenszemek vagy a spórák esetében. Ekkor tehát az új sejtfalrétegek kívülről épülnek az elsődleges sejtfalra, a sejt üregének mérete pedig nem változik. Ez a centrifugális sejtfalvastagodás (6. ábra).
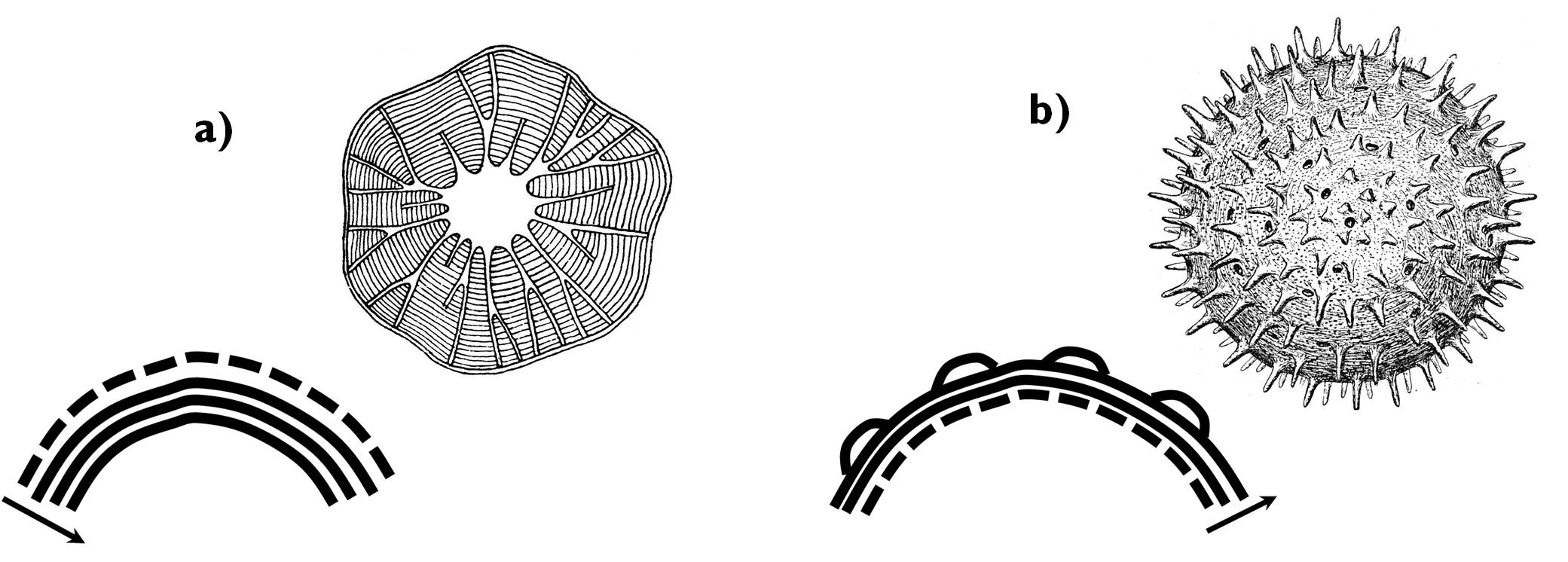
6. ábra. A centripetális (a) és a centrifugális (b) sejtfalvastagodás.
(Szaggatott vonal jelöli az elsődleges, míg folytonos a másodlagos sejtfalrétegeket.)
(a: Kárpáti nyomán, b: Sárkány és Szalai nyomán)
A sejt energiatermelő központjai: a mitokondrium és a színtest
Minden élő sejt energiát igényel életfolyamatainak fenntartásához. Ez az energia – az anyagcsere résztvevői számára hozzáférhető módon – a sejten belül kémiai kötések formájában, az adenozin-trifoszfát (ATP) nagy energiájú (makroerg) pirofoszfát kötéseként raktározódik. Intenzív ATP-szintézis az eukarióta sejt organellumai közül a mitokondriumban és a színtestben zajlik (a citoszólban csak kisebb arányban). A két sejtalkotó közül a mitokondriumok le is adják a sejtplazmába a bennük lebontó folyamatok kapcsán keletkező ATP nagy részét, hogy az egyéb, energiát igénylő folyamatokban hasznosulhasson. A színtestek ezzel szemben szerves anyagok szervetlen vegyületekből történő szintézisére fordítják a fényenergia segítségével előállított ATP-t.
A mitokondrium és a színtest evolúciós eredete
Amint arról már az eukarióta sejt kialakulásáról szóló fejezetben is szó volt, e két sejtalkotó evolúciós eredetét Lynn Margulis endoszimbionta elméletével magyarázzuk. Eszerint a mitokondriumok és a színtestek valaha önálló életet élt prokarióta sejtek leszármazottai: az előbbi a bíbor nem-kén baktériumok közé tartozó, hatékony aerob lebontásra (citromsav-ciklusra és terminális oxidációra) képes csoportból, míg a színtest ősi fotoszintetizáló kékalgákból alakult ki.
A mitokondrium és a színtest evolúciós eredete
A közös evolúciós múlt eredményeképpen a két sejtalkotó felépítésében és működésében számos közös vonás felismerhető. Mindkettőt kettős burkolómembrán határolja, amelyek közül a belső jóval nagyobb felületű, redőzött, és betüremkedik a sejtalkotó alapállományába, sőt akár (a színtest esetében) jellegzetes lefűződéseket is létrehozhat
Az ATP-szintézis mechanizmusa: a kemiozmotikus hipotézis
A mitokondriumok felépítése és működése
A mitokondriumok néhány mikrométer hosszú, gömbölyded, pálcika vagy fonál alakú sejtalkotók, amelyek minden olyan eukarióta sejtben előfordulnak, amelyekben aerob lebontási folyamat (biológiai oxidáció) végbemegy. Számuk a sejt energiaigényétől függően változik. Kettős borítómembránjuk belső tagja nagyméretű redőket, ún. krisztákat (christa) hoz létre az organellum belsejében (7. ábra). (A krisztás mitokondriumok mellett léteznek csöves belső szerkezetű, tubuláris mitokondriumok is, például számos gombacsoport sejtjeiben.) A mitokondrium alapállománya a mátrix. Itt találjuk a mitokondrium génjeit hordozó, gyűrű alakú DNS-molekulákat is (szinte mindig több példányban), valamint a fehérje-szintetizáló apparátust (riboszómákat, tRNS-eket, mRNS-eket).
A mitokondrium a biológiai oxidáció végső folyamatainak a színtere. A sejtplazmában végbemenő glikolízis termékei a mitokondrium mátrixában zajló citromsav-ciklusban alakulnak CO2-dá. Eközben a belső membránrendszerhez kötötten folyó terminális oxiodáció nagy mennyiségű ATP-t termel.
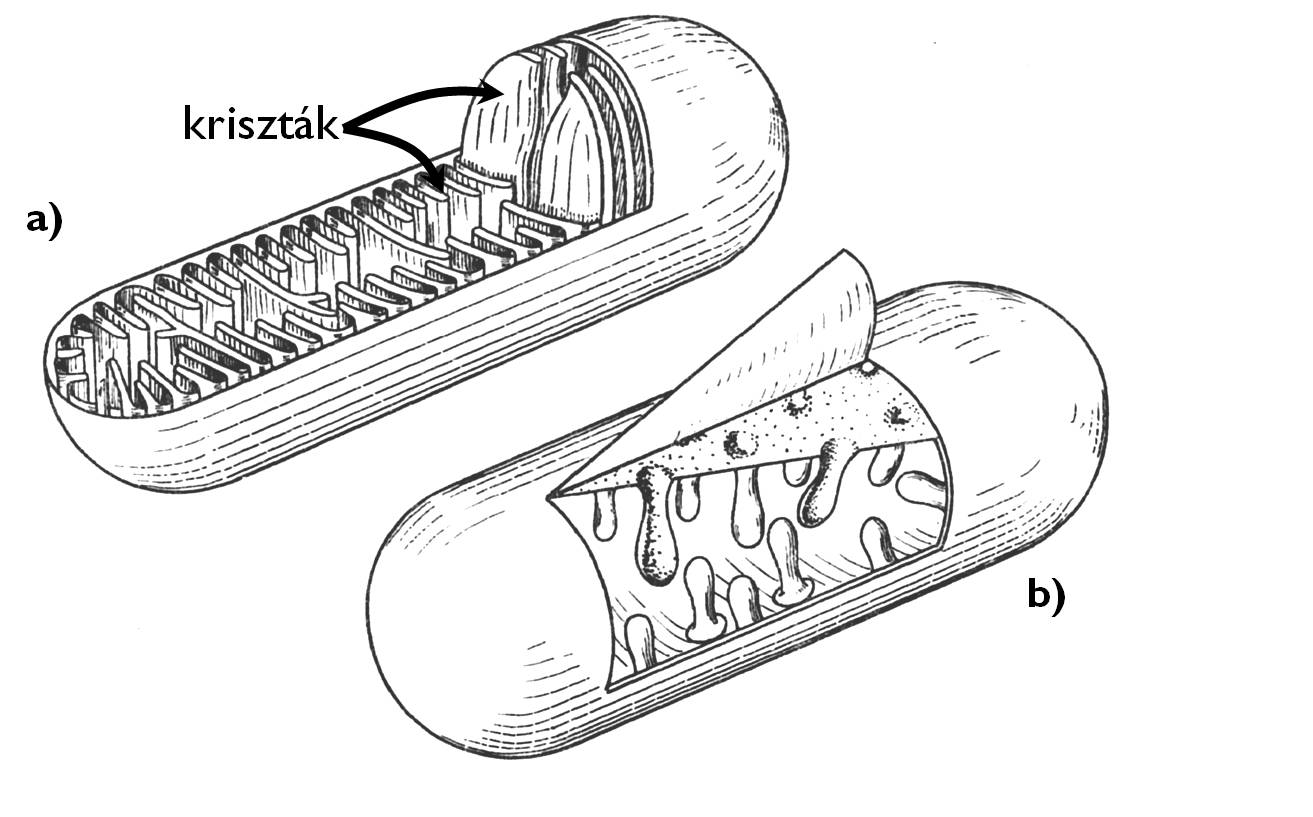
7. ábra. A krisztás (a) és a tubuláris (b) mitokondrium felépítése. (Haraszty nyomán)
A mitokondriumban zajló anyagcsere-folyamatok
A színtestek (plasztiszok) felépítése és működése
A színtestek változatos megjelenésű és méretű sejtalkotók. Számuk egy-egy növényi sejtben igen tág határok között változik: egyes algasejtek egyetlen, míg egy zárvatermő növény átlagos fotoszintetizáló sejtje 20-60 darabot tartalmaz belőlük. A színtestek (plasztiszok) általános felépítésére jellemző, hogy kettős burkolómembránjuk közül a belső többszörösen redőzött, s előbbiből lefűződő, keskenyebb vagy kiszélesedő, zárt ciszternák alkotta membránrendszert alkot; ez az ún. tilakoid. A színtestek alapállománya a sztróma, ami a mitokondrium mátrixához hasonlóan egyéb fehérjék mellett a sejtalkotó saját, gyűrű alakú DNS-ét, RNS-eket és a fehérjeszintézishez szükséges (prokarióta típusú) riboszómákat is tartalmazza.
A zöld színtest (kloroplasztisz)
A kloroplasztisz a növényi anyagcsere sajátosságát (foto-autotrófia) meghatározó folyamat, a fotoszintézis színtere. Nevét jellemző színéről kapta, ami a fotoszintézisben alapvető szerepet játszó pigmentmolekulák, a klorofillok jelenlétének köszönhető. Valamennyi zöld növényi szövetben, szervben előfordul (a levél, a fiatal szár, az éretlen termések, egyes takarólevelek klorenchima szöveteiben). A kloroplasztiszok ultrastrukturális jellemzője, hogy tilakoidmembránjuk adott helyeken egymásra helyezett korongokhoz hasonló rendszert, gránumokat hoz létre (8. ábra).
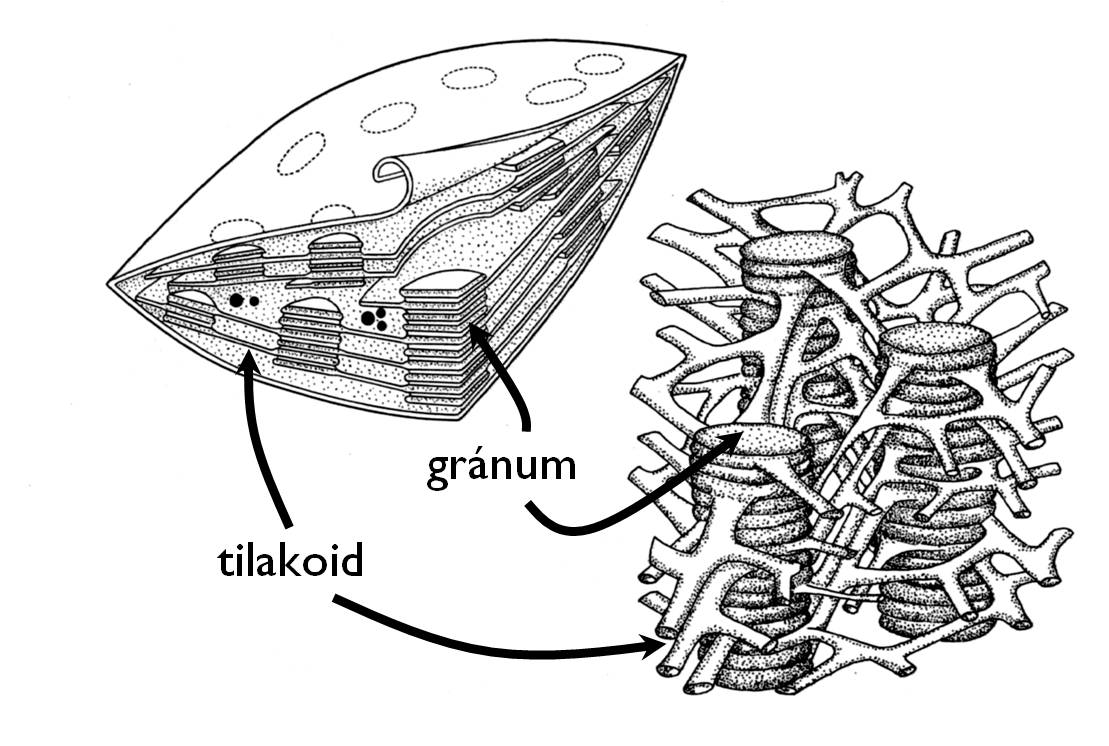
8. ábra. A kloroplasztisz felépítése. (Haraszty nyomán)
A kloroplasztiszban zajló anyagcsere-folyamatok
Egyéb színtesttípusok
Az osztódószövetek sejtjeinek színtestei még differenciálatlanok, bennük a tilakoidmembrán kis felületű, gránumokat nem alkot. Ezek az egyszerű felépítésű színtestek a proplasztiszok, amelyekből a színtestek összes többi típusa kialakulhat. A proplasztiszokra jellemző az osztódás (hasadás) képessége is.
Raktározó funkciót látnak el a különféle színtelen színtestek (leukoplasztiszok). Ezekben felhalmozódhat fehérje ( proteinoplasztisz), olaj (elaioplasztisz), de a leggyakoribb tartaléktápanyag az amiloplasztiszokban tárolt keményítő. Valamennyi leukoplasztisz-típusra jellemző, hogy bennük a tartalékanyagokat kettős membrán veszi körül (a színtest burokmembránjai), és hogy tilakoid-membránjuk erősen visszafejlődik. Fehérjét tartalmaznak a Commelinaceae család /komelinafélék/ tagjainak (pl. Zebrina pendula /zebrapletyka/) levelének epidermiszsejtjei, elaioplasztiszok főleg magvakban és termések falában fordulnak elő (9. ábra).
A keményítőszemcsék sokfélesége
A színtestek egy következő típusa a színes színtest (kromoplasztisz). Ezek nagy mennyiségű karotinoid típusú vegyületet raktároznak, amelyek többnyire kristályos formában válnak ki a színtest belsejében, ezzel a kristálynak megfelelő alakúvá torzítva azokat. A kromoplasztiszok a narancsos–vöröses színű, karotinoidokat raktározó növényi részek (egyes raktározó szervek, termések, virágok) színét adják (10. ábra).
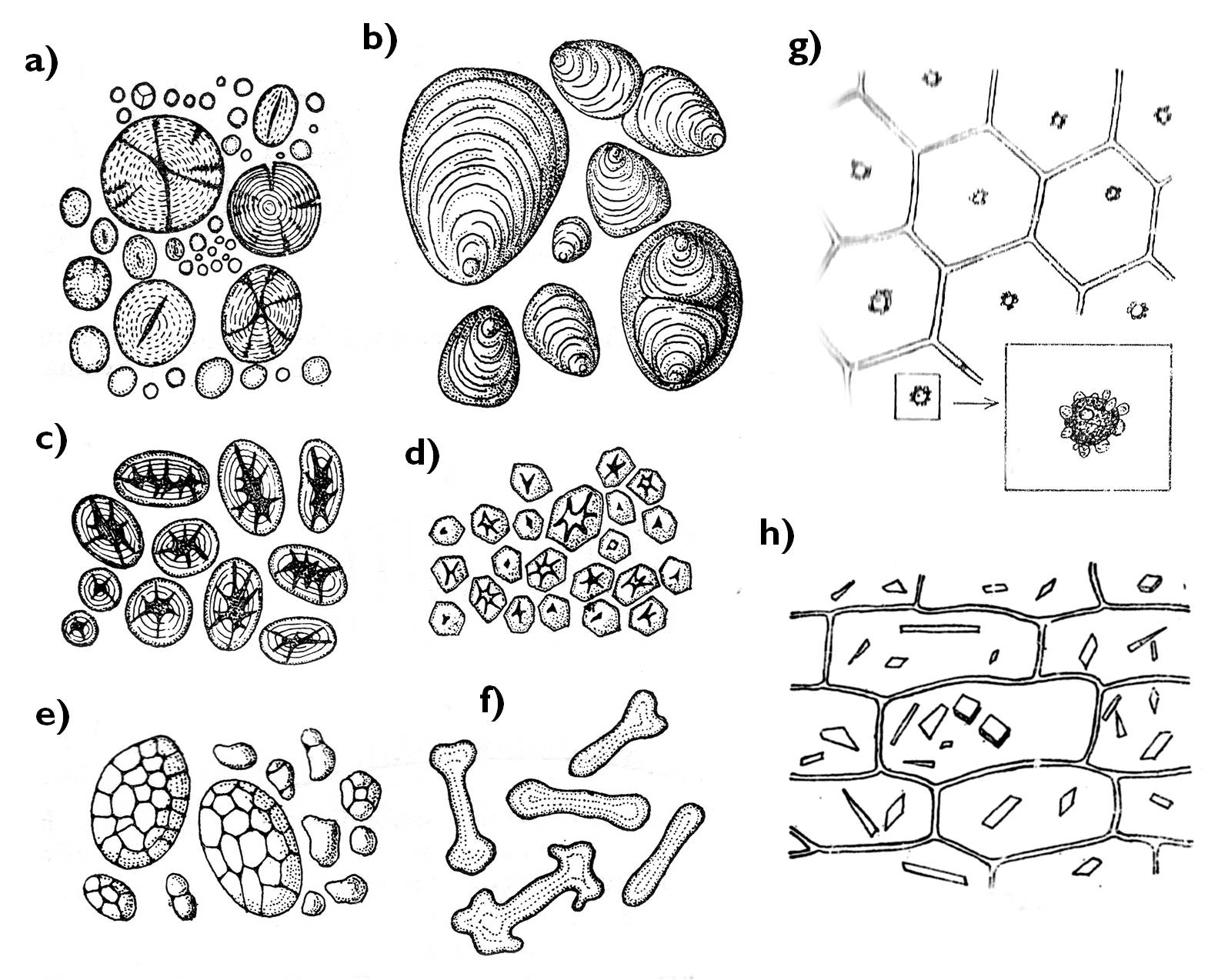
9. ábra. Színtesttípusok.
a)-f) amiloplasztisz (a: Triticum, b: Solanum, c: Phaseolus, d: Zea, e: Avena, f: Euphorbia); g: proteinoplasztisz (Zebrina), h: kromoplasztisz (Daucus).
(a-f: Kárpáti nyomán, g-h: Sárkány és Szalai nyomán)
Egy különleges színtest: az etioplasztisz
A színtestek egymásba alakulása
A növényi sejt metabolikus kapcsolatrendszere (az alapvető biokémiai folyamatok lokalizációja a sejtben)
A sejtmag felépítése és működése
A sejtmag felépítése
Az eukarióta sejtek legfontosabb, a prokariótáktól megkülönböztető sejtalkotója a sejtmag. Örökítőanyaguk nem a sejtplazmában szabadon, hanem attól egy kettős membránnal (sejtmaghártya) elkülönítve helyezkedik el. A sejtmaghártya az evolúció során nagy valószínűséggel a DER-ból jött létre. A két membrán-sejtalkotó szoros kapcsolata még ma is egyértelmű: a DER membránrendszere folytonosan megy át a sejtmaghártyába. Emellett a két organellum felépítése is igen hasonló: mindkettő felületén riboszómák ülnek, membránjaikat hasonló lipidkomponensek építik fel.
Az örökítőanyag szerveződése
A DNS kettős spirálja igen vékony (2 nm), ezért érzékeny, törékeny molekula. Mivel a DNS-ben hordozott információt a nukleotidok sorrendje adja (hasonlóan ahhoz, ahogyan az írott szöveg információját a betűk sorrendje kódolja), minden a DNS-szerkezetet érintő sérülés információvesztést okoz. Ennek elkerülésére a DNS nem szabadon úszik a magplazmában, hanem hisztonfehérjékből álló korongokra tekerve helyezkedik el. A hisztonkorong és a rátekert DNS egysége a nukleoszóma. A nukleoszomális szerkezet mindig megfigyelhető a sejtmagban, a DNS még a transzkripció időszakára sem tekeredik le a fehérjekorongról (az átírást végző enzim „beférkőzik” a hisztonfehérjék és a DNS közé).
A DNS legjobban feltekert formája a sejtosztódás alatt megfigyelhető, jellegzetes alakú kromoszóma. A kromoszómák kialakulása során a hisztonkorongokra tekert DNS a kromoszóma vázát alkotó szerkezeti fehérjékhez („scaffold-proteinek”) kapcsolódik. A kromoszómák kialakulásának előnye, hogy a DNS-molekulák sérülése nélkül lehet mozgatni, elosztani az örökítőanyagot a sejtosztódás során az utódsejtek között (lásd később).
A kromoszómák látható formájukban (azaz a sejtosztódás alatt) többnyire két félből, ún. kromatidából állnak. (A kromatidák letekeredett, tehát sejtosztódás közti állapotukban is egymás közelében vannak. A sejt ezen életszakaszában előfordulhatnak azonban egy kromatidás kromoszómák is. – Részletesen lásd alább!) A két kromatidát az ún. elsődleges befűződés (centromeron) kapcsolja össze (emellett a kromoszómán lehetnek további – másodlagos, harmadlagos – befűződések is). A kromatidák centromerontól a kromoszómáig tartó szakasza az ún. kromoszómakar. A kromoszómakarok végén ismétlődő DNS-szekvenciákból álló ún. telomer régió helyezkedik el, amelynek az a feladata, hogy védje az információt (géneket ill. szabályzó régiókat) kódoló DNS-szakaszokat a sérülésektől, letöredezéstől.
Egy kromatidát mindig egyetlen DNS-molekula alkot. A kétkromatidás kromoszómákban tehát két darab DNS-kettős spirál helyezkedik el. A kétkromatidás kromoszómák DNS-molekuláinak információtartalma (azaz nukleotidsorrendje) tökéletesen azonos. (Ennek az a magyarázata, hogy az egy kromatidás kromoszómákból csak a sejtciklus adott fázisában, a DNS-másolás (replikáció) eredményeképpen kapott, azonos DNS-molekulák összekapcsolódásával jönnek létre a két kromatidás kromoszómák.)
A kromoszómákon találjuk a géneket. A gén az adott tulajdonságot (azaz pontosabban egy adott fehérjét vagy RNS-t) meghatározó DNS-szakasz megnevezése. A gén helye a kromoszómán a lókusz. Egy adott tulajdonság (pl. pollenalak, virágszín) nem csak egyféle formában fordulhat elő a fajon belül (létezhetnek pl. hosszúkás és kerekded pollenű, fehér és piros virágszínű egyedei is a fajnak). Ebben az esetben a génnek is több változata, allélje fordul elő. A sejtben található gének összességét hívjuk a sejt (azaz az élőlény) genomjának. Fontos megjegyezni, hogy eukarióta sejtek esetében a teljes genomnak csak egy része található a sejtmagban (nukleáris genom), hiszen emellett DNS-t találunk egyes sejtalkotókban (mitokondrium, színtest) is (ez az ún. extranukleáris genom).
A gének illetve allélek lokalizációjáról tehát elmondhatjuk, hogy egy adott kromoszómának a két kromatidáján azonos lókuszban azonos gének, szükségszerűen azonos alléljei vannak jelen (a két kromatidát alkotó DNS-ek azonossága miatt). Az egy kromoszómán elhelyezkedő gének ún. kapcsoltsági csoportot alkotnak, ami azt jelenti, hogy – kromoszomális sérülésektől eltekintve – a faj minden egyedében egy adott kromoszómán mindig ugyanazon tulajdonságok génjei helyezkednek el (legfeljebb az allélek térnek el egymástól).
Amennyiben a hajtásos növények egyetlen testi sejtjét megvizsgáljuk, abban az egy kromoszómán levő (azaz egy kapcsoltsági csoportot alkotó) gének nem egy, hanem két változatban fordulnak elő, mivel ezen sejtek sejtmagjában minden kromoszóma két példányban, ún. homológ kromoszómapárok formájában van jelen. A homológ kromoszómákon ugyanazok a gének fordulnak elő, de nem feltétlenül azonos allélek formájában. A kromoszómapár tagjai eredetükben különböznek egymástól: az egyik kromoszóma a petesejtből, a másik a hímivarsejtből származik a megtermékenyítés folyamatában. Azokat a sejteket, amelyekben a kromoszómák csak egy példányban vannak jelen, haploid (egyszeres információtartalmú) sejteknek nevezzük, míg a kromoszómapárokat tartalmazó sejtek a diploid (kétszeres információtartalmú) sejtek. Az élővilágban az ivarsejtek mindig haploidok, míg a megtermékenyítés során ezek összeolvadásával létrejövő zigóta diploid. A növények körében az is előfordul, hogy három, négy vagy akár még több változatban is előfordulnak a kromoszómák (azaz egy kapcsoltsági csoport génjei), ilyenkor triploid, tetraploid illetve általában poliploid sejtekről beszélünk.
A sejtosztódás
A többsejtű növényi szervezet kialakulása során, valamint a növényi életciklus stádiumai közti átmenet részeként az eukarióta sejteknek többszöröződniük, osztódniuk kell. Ennek a folyamatnak szükségszerű része a sejtmagban található információ, azaz a DNS szétosztása az utódsejtek között. Ez csak úgy történhet meg, ha az igen hosszú, vékony, törékeny DNS-molekulák szorosan feltekerednek, azaz kromoszómákká alakulnak. A sejtosztódást megkezdő sejtekben mindig két kromatidás kromoszómákat találunk. A sejtosztódás valamely szakaszában a kromoszómákhoz speciális mikrotubulusok, ún. húzófonalak kapcsolódnak (ezek összessége a magorsó), amelyek szétválasztják egymástól az egyes kromatidákat, ezzel kettéosztva a kiindulási sejtmagban levő DNS-t. (Az állati sejtek esetében a húzófonalak a két, egymásra merőleges henger alakjába rendeződő mikrotubulusok alkotta sejtközpont körüli régióból indulnak ki. A növényi sejtek többségéből ez a sejtalkotó hiányzik.)
A sejtosztódással kialakuló utódsejtek közti sejtfal, azaz az osztódási sík helye már viszonylag korán, az osztódás korai szakaszában eldől (lásd később). Amennyiben ez a kiindulási sejt középvonalában található, tehát két egyforma sejt jön létre, ekvális sejtosztódásról, ellenkező esetben inekvális osztódásról beszélünk. (Inekvális osztódás vezeti be valamennyi speciális sejt, egyszerű szövet kialakulását, például a növényi szőrök differenciálódását a protoderma-sejtekből, de inekvális a zigóta első osztódása is – lásd a 4. fejezetben!) A sejtosztódás jellemzője az is, hogy az újonnan kialakuló sejtfal párhuzamos-e az adott szerv felszínével vagy merőleges arra. Az első esetben az osztódást periklinálisnak, az utóbbi esetben antiklinálisnak nevezzük.
A mitózis (számtartó osztódás)
A mitózis során – magyar nevének megfelelően – anélkül történik meg a DNS szétosztása az utódsejtek között, hogy megváltozna a kromoszómaszám, azaz a ploidiaszint. (Haploid kiindulási sejtből két darab haploid, diploid kiindulási sejtből két darab diploid sejt keletkezik stb.) Ez úgy képzelhető el, hogy a folyamatban a két kromatidás kromoszómák testvérkromatidái válnak ketté, és jutnak – külön-külön – a két utódsejtbe (10. ábra). Mivel a testvér-kromatidák DNS-ének nukleotidsorrendje azonos, a mitózissal keletkező utódsejtek információtartalma is azonos, és megegyezik a kiindulási sejt információtartalmával (tehát nem történik a meiózisra jellemző genetikai rekombináció).
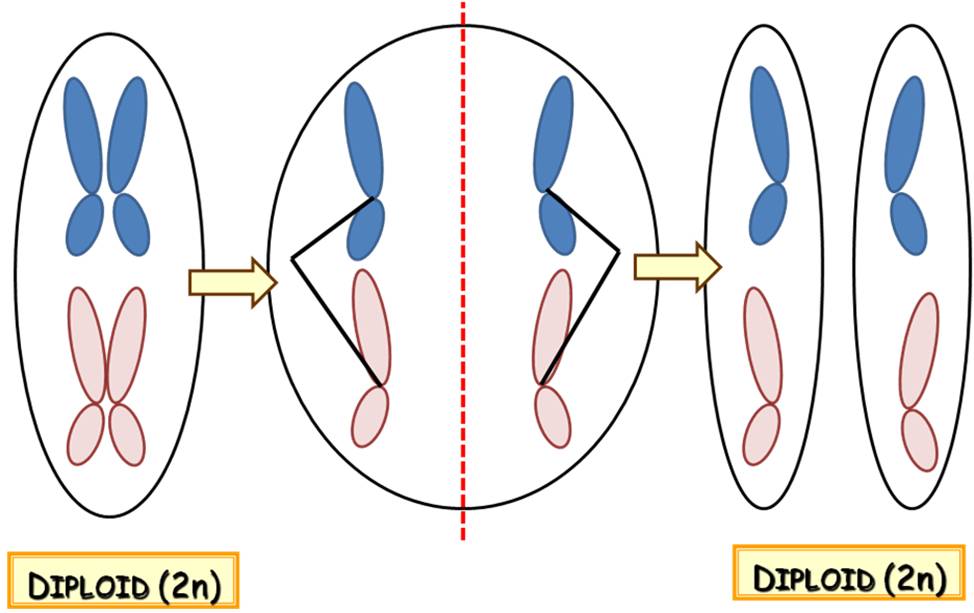
10. Animáció a mitózis folyamatáról : a kromatidák mozgását és szétválását mutatja be.
Mitózissal jönnek létre a növények testi sejtjei (függetlenül attól, hogy a haploid gametofiton, vagy a diploid sporofiton szakaszhoz tartozó képletekről van-e szó – lásd később!), valamint a növényi ivarsejtek is.
A meiózis (számfelező osztódás)
A számfelező sejtosztódás (meiózis) nevét onnan kapta, hogy folyamata során megfeleződik a sejtben levő kromoszómaszám, tehát a homológ kromoszómapárok tagjai is különböző utódsejtekbe kerülnek, nem csak a testvérkromatidák válnak szét egymástól. A folyamat két lépésben megy végbe (11. ábra). Az első főszakaszban a homológ kromoszómapárok elkülönülése történik meg, tehát a diploid, két kromatidás kromoszómákat tartalmazó sejtből két darab, immáron haploid, de még mindig két kromatidás kromoszómákat tartalmazó sejt jön létre. A meiózis második főszakaszában a haploid sejtekben levő kromoszómák kromatidái válnak szét. A számfelező osztódás eredményeképpen tehát összesen négy darab, haploid, egy kromatidás kromoszómákat tartalmazó utódsejt jön létre.
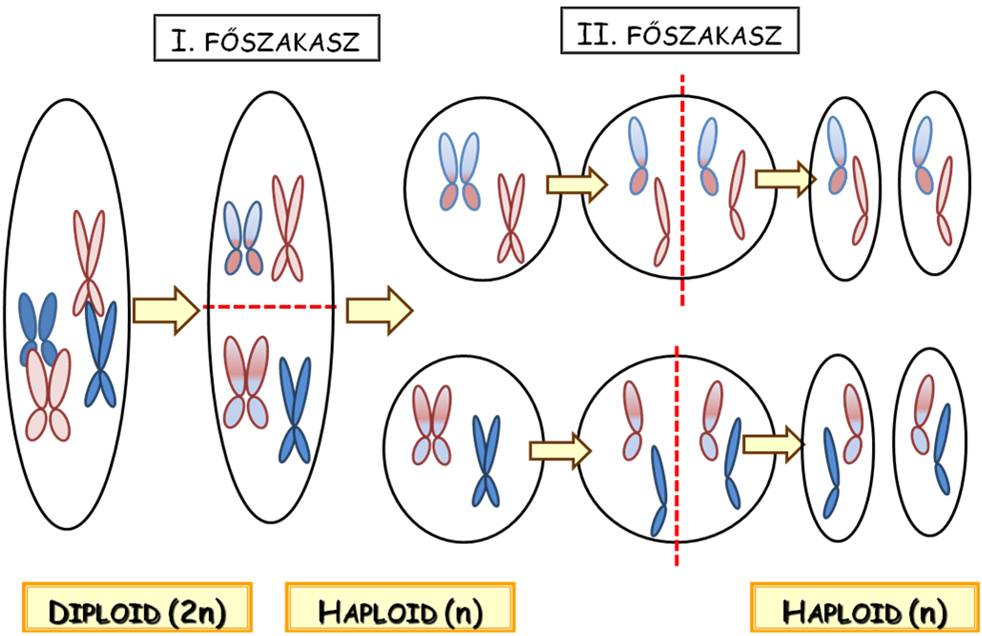
11. Animáció a meiózis folyamatáról : a kromoszómák és a kromatidák mozgását és szétválását mutatja be.
Mivel a meiózis során a homológ kromoszómapárok tagjai különböző utódsejtekbe kerülnek, a keletkező haploid sejtek genetikai információtartalma bizonyosan különbözni fog egymástól. Mivel teljesen véletlenszerű, hogy az egyes kromoszómapárok apai vagy anyai tagja vándorol-e a sejt adott pólusa irányába (az első főszakasz anafázisában), az utódsejtek a szülői allélkombinációkhoz képest új összetételben tartalmazzák majd a haploid sejtet képző egyed szüleitől származó alléleket, azaz genetikailag rekombinánsok lesznek. A meiózisnak éppen az az elsődleges célja, hogy a genetikai rekombináció révén biztosítsa a genetikai sokféleséget.
A számfelező osztódás során nem csak a különböző géneket hordozó kromoszómák alléljeinek szintjén mehet végbe rekombináció. Lehetőség van az egyazon kromoszómán levő lókusszal rendelkező gének alléljeinek keveredésére is. Az átkereszteződés (crossing over) során, a meiózis első profázisában ugyanis a párba álló homológ kromoszómák egyes szakaszai (azaz a megfelelő DNS-szakaszokon levő allélek) kicserélődhetnek egymással. Ekkor tehát az apai kromoszómának egy kis darabja „helyet cserél” az anyai kromoszóma megfelelő darabjával, s így már anyai, és apai eredetű allélok egyaránt lesznek egyazon kromatidán.
Meiózissal a növényvilágban az életfázis haploid (gametofiton) szakaszának első sejtjei jönnek létre: a spórák. (Itt még egyszer emlékeztetnénk rá, hogy a növényi ivarsejtek a gametofiton szakasz utolsó sejtjei, haploid sejtek osztódásával alakulnak ki, tehát szükségszerűen mitózissal keletkeznek!)
4. táblázat. A mitózis és a meiózis öszehasonlítása.
|
Mitózis |
Meiózis |
|
Számtartó osztódás |
Számfelező osztódás |
|
A ploidiaszint nem változik |
A kromoszómák száma megfeleződik |
|
Az utódsejtek genetikailag azonosak egymással |
Az utódsejtek genetikailag különböznek egymástól és a szülői sejttől is |
|
2 db utódsejtet eredményez |
4 db utódsejtet eredményez |
|
A DNS-szétválasztása egy lépésben történik: a testvérkromatidák szétválása |
A DNS-szétválasztása két lépésben történik: I. főszakasz: homológ párok szétválása; II. főszakasz: a testvérkromatidák szétválása |
|
Szakaszai: |
Szakaszai: |
|
Így keletkeznek a testi sejtek, ivarsejtek |
Így keletkeznek a spórák |
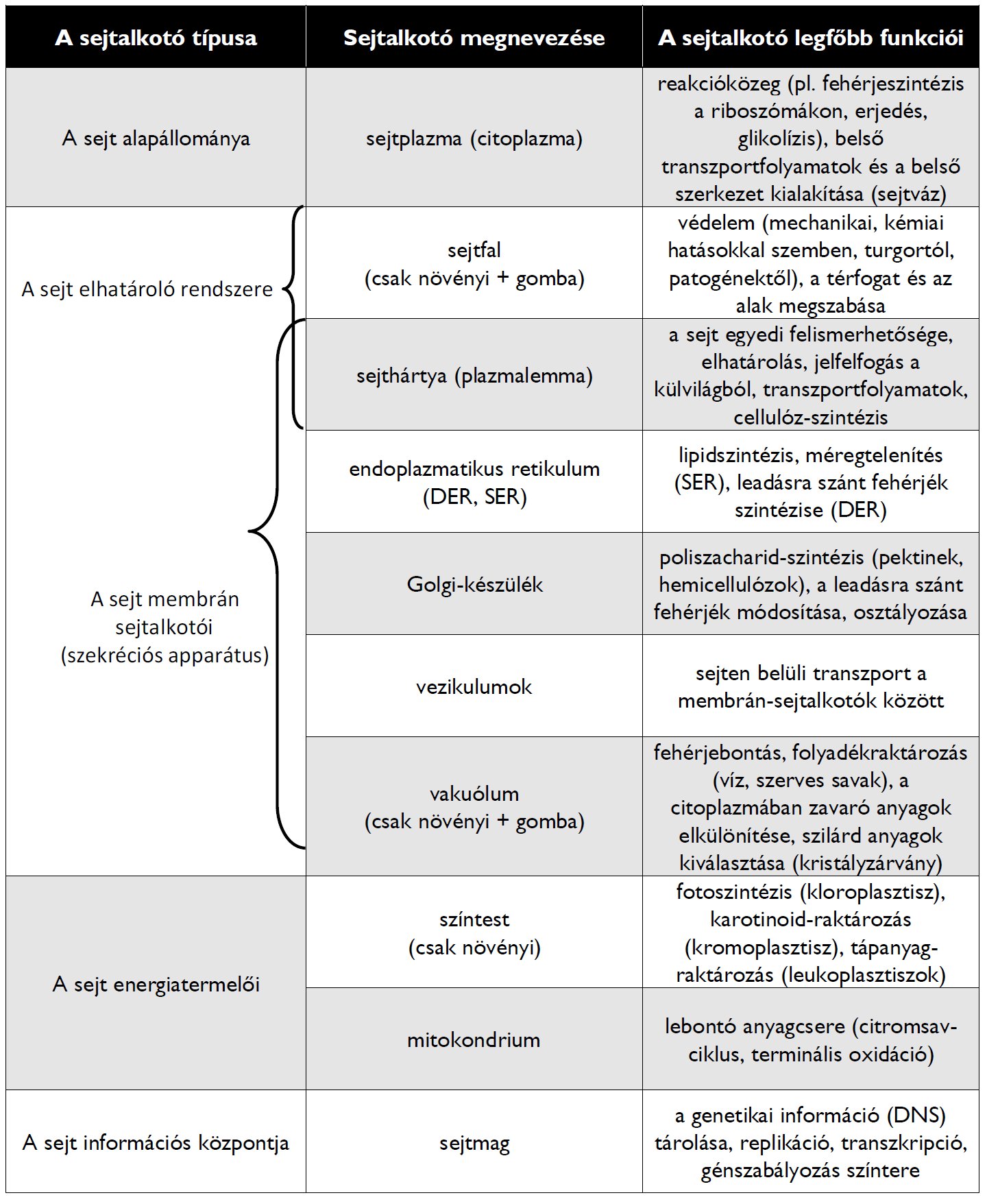
5 . táblázat. A legfontosabb eukarióta sejtalkotók áttekintése.
Ellenőrző feladatok
Ide jó lenne a feladatokat feltölteni
Irodalomjegyzék
Beck CB (2010): An Introduction to Plant Structure and Development: Plant Anatomy for the Twenty-First Century. 2. kiad. Cambridge University Press, Cambridge,
Darnell J, Lodish H, Baltimore D (szerk.) (1986): Molecular Cell Biology. Revised Printing with Expanded Index. Scientific American Books, New York.
Evert RF, Eichhorn SE (szerk.) (2006): Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body – Their Structure, Function, and Development, 3. kiad., Wiley-Interscience, New Jersey.
Gyurján I (1996): Növényszervezettan. Egyetemi jegyzet. ELTE Eötvös K., Bp.
Haraszty Á (1998): Növényszervezettan és növényélettan. 6. kiad., Nemzeti Tankvk., Bp.
Jacob F, Jäger EI, Ohmann E (1985): Botanikai kompendium. Natura, Bp.
Kovács János (1999): Sejttan. – In: Sass Miklós, Zbory Géze (szerk.): Összehasonlító anatómiai előadások I. ELTE Eötvös Kiadó, Bp.
Láng F (szerk.) (1998): Növényélettan. A növényi anyagcsere. ELTE Eötvös K., Bp.
Mauseth JD (2008): Plant Anatomy. The Blackburn Press, New Jersey.
Sárkány S, Szalai I (1966): Növénytani praktikum I. Növényszervezettani gyakorlatok. 3. kiad., Tankvk., Bp.
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028