Növényi szaporodásbiológia
Szerző: Erős-Honti Zsolt
Az ivaros és az ivartalan szaporodás
Az élővilág egyéb csoportjaihoz hasonlóan a szaporodási folyamatoknak a növények körében is két célja lehet, az egyedszám gyarapítása vagy a genetikai sokféleség biztosítása. Az elsődlegesen az egyedszám növelését célzó, genetikai állandósággal járó szaporodási módokat ivartalan, míg a genetikai változékonyságot létrehozó folyamatokat ivaros szaporodásnak nevezzük (1. táblázat).
Az ivartalan szaporodás sajátossága, hogy nem jár a genetikai információ megváltozásával (rekombinációval), tehát az utódok (jelen esetben klónok) génállománya megegyezik és a szülő egyedével is azonos. E szaporodási folyamatok során nem alakulnak ki ivarsejtek, és a rekombinációt biztosító osztódási módra, a meiózisra, sem kerül sor. Az ivartalan szaporodás történhet mitózissal létrejövő szaporítósejtek (mitospórák) segítségével, a vegetatív test feldarabolódásával kialakuló egységekkel (pl. egy mohapárna darabjai) vagy vegetatív szervek módosulataival (pl. tarack, inda) is. Mivel ezen folyamatok során a génállomány nem változik, az adott növényfaj populációja a környezeti változásokhoz nem tud gyorsan alkalmazkodni, így az ivartalan szaporodás elsősorban a többé-kevésbé állandó környezeti körülmények között előnyös.
Az ivaros szaporodás lényege a genetikai információ rekombináció révén történő megváltozása, a genetikai változékonyság biztosítása. A folyamat során – a többsejtű élőlények körében – meiózis is történik, valamint ivarsejtek kialakulására és ezek összeolvadására (megtermékenyítésre) is sor kerül. Az ivaros szaporodás szervei a generatív szervek (növénycsoporttól függően az ivarszervek, a virág, a mag, a termés). A keletkezett utódok száma messze elmaradhat az ivartalan szaporodás során megfigyelhető utódszámtól, az utódok génállománya azonban mind egymástól, mind a szülői egyedekétől eltérő. Természetes populációkban a folyamat elsődleges célja a genetikai változékonyság biztosítása révén a változó környezethez történő alkalmazkodás.
1. táblázat. Az ivartalan és az ivaros szaporodás összehasonlítása.
|
Ivartalan szaporodás |
Ivaros szaporodás |
|
|
A folyamat elsődleges célja |
az egyedszám növelése |
genetikai sokféleség biztosítása |
|
Rekombináció |
nincs |
van |
|
Utódok génállománya |
azonos egymással, szülőkkel (klónok) |
különböznek egymástól, szülőktől |
|
Alapfolyamatai |
mitózisok sorozata |
meiózis, ivarsejtképzés, megtermékenyítés |
|
Szaporító képletei |
vegetatív szervek (módosulatok) |
generatív szervek |
|
Milyen környezetben előnyös? |
közel állandó körülmények |
gyorsan változó körülmények |
Az ivartalan és ivaros szaporítás
A növényi szaporodás általános vonásai, evolúciója
A növényi szaporítósejtek
A növények szaporodási folyamataiban kétféle szaporítósejt vesz részt: az ivarsejtek (gaméta) illetve a spórák. Az ivarsejtek olyan haploid sejtek, amelyek a megtermékenyítés folyamatában összeolvadva az életciklus első diploid sejtjét, a zigótát hozzák létre. Az ivarsejtek lehetnek mozgásra képes zoogaméták (planogaméták) vagy mozdulatlan aplanogaméták. A szárazföldi növények csoportjainál egyértelműen megkülönböztethetők a hímivarsejtek a petesejtektől. Az előbbiek (spermatocita) kisebb méretű, nagyobb számban termelődő ivarsejtek, míg a petesejtek (oocita) nagyobbak, mozdulatlanok és jóval kevesebb termelődik belőlük. Az ivarsejtek a növények egyes csoportjaiban a hímivarszervben (antheridium) ill. a női ivarszervben (archegonium) termelődnek.
A spórák vastag fallal körülvett szaporítósejtek, amelyek sok esetben a kedvezőtlen időszak átvészelésére szolgáló kitartóképletek illetve a terjedést szolgáló propagulumok is egyben. Ősibb megjelenésű növénycsoportokban (algák körében) előfordulnak ostoros mozgásra képes zoospórák, de sokkal gyakoribbak a mozgásra képtelen ún. aplanospórák. A növényvilágban a spórák szinte minden esetben meiózissal kialakuló ún. meiospórák, de az algák körében előfordulnak az ivartalan szaporodást szolgáló, számtartó osztódással létrejövő mitospórák is. (Fontos megjegyezni, hogy a meiospórák nem azonosak az ivarsejtekkel. Ez utóbbiak ugyanis – a növények körében – mitózissal kialakuló haploid sejtek, amelyek a megtermékenyítés eseménye során zigótává olvadnak össze. A meiospórák ezzel szemben mitotikus osztódásokkal a haploid sejtekből áló gametofiton testet hozzák létre. Részletesebben lásd később!)
A növénycsoportok spóratípusai különböznek abból a szempontból, hogy egy adott meiospórából kialakuló növényi testen hím- és női ivarsejtek egyaránt megjelennek-e:
-
- Az izospórák esetében valamennyi spórából olyan növényi rész hajt ki, ami hím- és női ivarsejtek létrehozására egyaránt képes (azaz a spórák ivarilag nem elkötelezettek). Ilyen spórái vannak a lombosmohák egyes képviselőinek vagy a szárazföldi páfrányoknak.
- Az anizospórák ivarilag már elkötelezettek: vannak hím illetve női jellegű spórák, így a belőlük létrejövő testen csak a megfelelő ivarsejt termelődik.
-
- A homoiospórák esetében az ivarilag elkötelezett spórák alakilag nem különböztethetők meg egymástól (pl. a májmohák vagy a zsurlók esetében).
- A heterospórás növények (pl. vízi páfrányok, nyitva- és zárvatermők) hím jellegű spórái jóval kisebbek és nagyobb számban termelődnek, mint a női jellegű spórák. Ez utóbbi esetben az alaki eltérésnek megfelelően mikrospóráról (hím) és makrospóráról (női) beszélünk.
-
A növényi életciklus általános jellemzői
A növények országába tartozó fajok életciklusa során haploid és diploid sejtekből álló szakaszok váltakoznak egymással. Kivétel nélkül igaz, hogy az életciklus során egy adott ponton haploid ivarsejtek jönnek létre, amelyek a megtermékenyítés folyamatában a zigótát hozzák létre. Ettől eltekintve azonban – elsősorban az algacsoportok körében – többféle életmenetet különböztethetünk meg a különböző ploidiaszintű (magfázisú) életszakaszok aránya ill. felépítése alapján.
Diploid és haploid sejtek az életciklusokban: az életmenet-típusok
A szárazföldi növények (mohák, harasztok, nyitva- és zárvatermők) ún. haplodiplonta, heteromorf életmenete két szakaszra, a zigótától a spóra-anyasejtekig tartó diploid (2n) sporofiton szakaszra és a spóráktól az ivarsejtek kialakulásáig tartó haploid (1n) gametofiton szakaszokra osztható (1. ábra). A meiózissal kialakuló spórákból egy egyszerűbb felépítésű test, az ún. előtelep jön létre, később ezen (vagy az ebből kialakuló összetettebb testen) mitózissal alakulnak ki az ivarsejtek. Az ivarsejtek összeolvadása visz át a sporofiton szakaszba, ahol a zigótából minden esetben embrió fejlődik, majd ennek további differenciálódásával alakul ki a növény diploid teste (sporofitonja). A sporofiton testen létrejövő spóraanyasejtek meiózisa lesz az az esemény, amivel kezdetét veszi az újabb gametofiton szakasz.
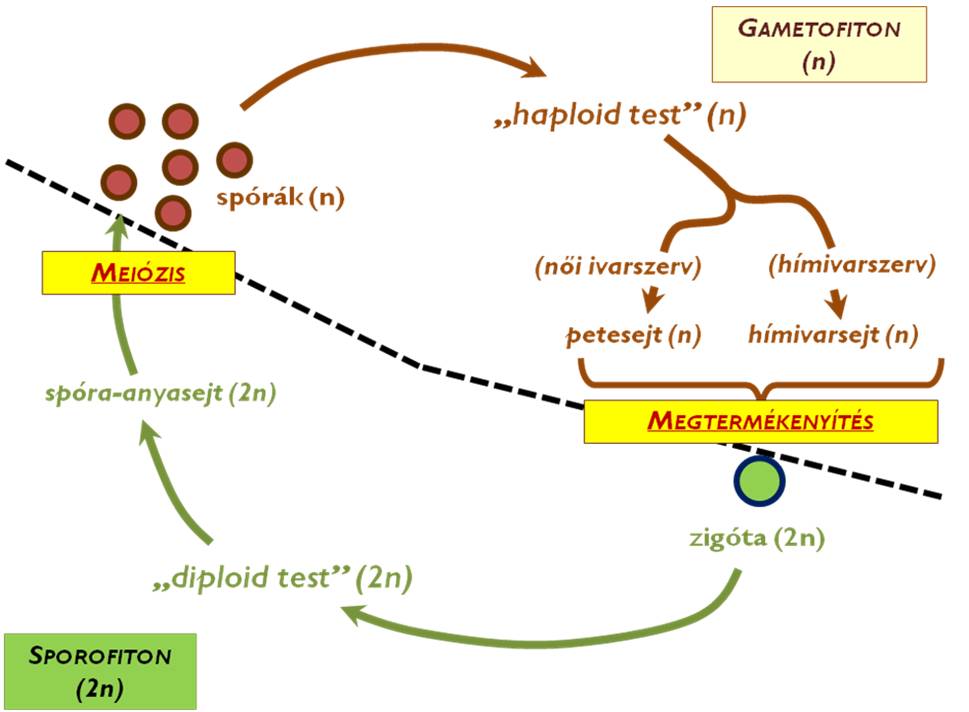
1. ábra. Az általános növényi életciklus.
A szárazföldi növények szaporodásának evolúciója
Az ősi zöldalgák igen változatos életmenettel rendelkező csoportjaiból két vonalon indult meg a szárazföldre lépő növénycsoportok evolúciója: az egyik ágon a mohák, míg a másikon a hajtásos növények (ősharasztok) első képviselői jelentek meg. A későbbi időszakok során az előbbi vonal oldalágnak bizonyult. Csak az ősi harasztok leszármazottaiból fejlődtek ki összetettebb testfelépítésű növénycsoportok (a nyitvatermők valamint a zárvatermők). A két vonal közös, az ősi zöldalgáktól eltérő szaporodásbiológiai jellemzője, hogy zigótájukból nem közvetlenül a diploid test fejlődik ki, hanem egy egyszerűbb felépítésű embrió, ami a későbbi differenciáció folyamatában válik majd összetettebbé. (Ezért is nevezik összefoglaló néven a szárazföldi növényeket Embryophyta-nak.)
A hasonlóságok ellenére a két evolúciós ág életciklusa között alapvető különbségek vannak. Ezek közül a legfontosabb a gametofiton és a sporofiton szakaszok arányaiban mutatkozik meg: míg a mohák körében a haploid gametofiton szakasz dominanciája figyelhető meg (a fotoszintetizáló mohanövényke maga is haploid sejtekből áll), az edényes növények fotoszintetizáló szervezete kivétel nélkül a diploid sporofiton szakasz része, így ennek a szakasznak az aránya (mind időtartamát, mind összetettségét és méretét tekintve) meghaladja a gametofitonét. Az eltérésből fakadó különbség, hogy míg a mohanövényeken ivarsejtek jönnek létre, addig a hajtásos növények körében az ivarsejtek az előtelepen (vagy annak evolúciós származékain) jönnek létre, a növény a spórák kialakulásának a helye.
A harasztok szaporodása
Az első harasztokig vezető evolúciós útvonalon – az alapvető testfelépítési változások mellett – olyan fontos szaporodásbiológiai újítások is kialakultak, amelyek nemcsak az ősi zöldmoszatokétól, de a mohák jellemzőitől is különböznek. Az életciklusban a sporofiton szakasz válik meghatározóvá, hiszen a fotoszintetizáló növényt is diploid sejtek építik fel. A sporofiton testen a spórák kialakulása valódi sporangiumokban történik. Ezek módosult levelek (sporofillumok) tövében elhelyezkedő, jellegzetes mechanizmussal felnyíló spóratartó képletek.
A spórákból valamennyi harasztcsoportban előtelep hajt ki, ez azonban nem fonalas, hanem telepes (álszövetes) testszerveződési szintet képviselő protallium (2. ábra). Az anteridium és az archegónium ezen az előtelepen jön létre. A mitózissal kialakuló ivarsejtek összeolvadásához itt is vízcseppre van szükség, s a kialakuló zigóta (a sporofiton első sejtje) itt is az archegónium belsejében fejlődik embrióvá. Az embrióból kialakuló diploid növény azonban jelentősen túlnövi a protalliumot, ami később elsorvad: a sporofiton élettani szempontból is függetlenné válik a gametofiton szakasz sejtjeitől.
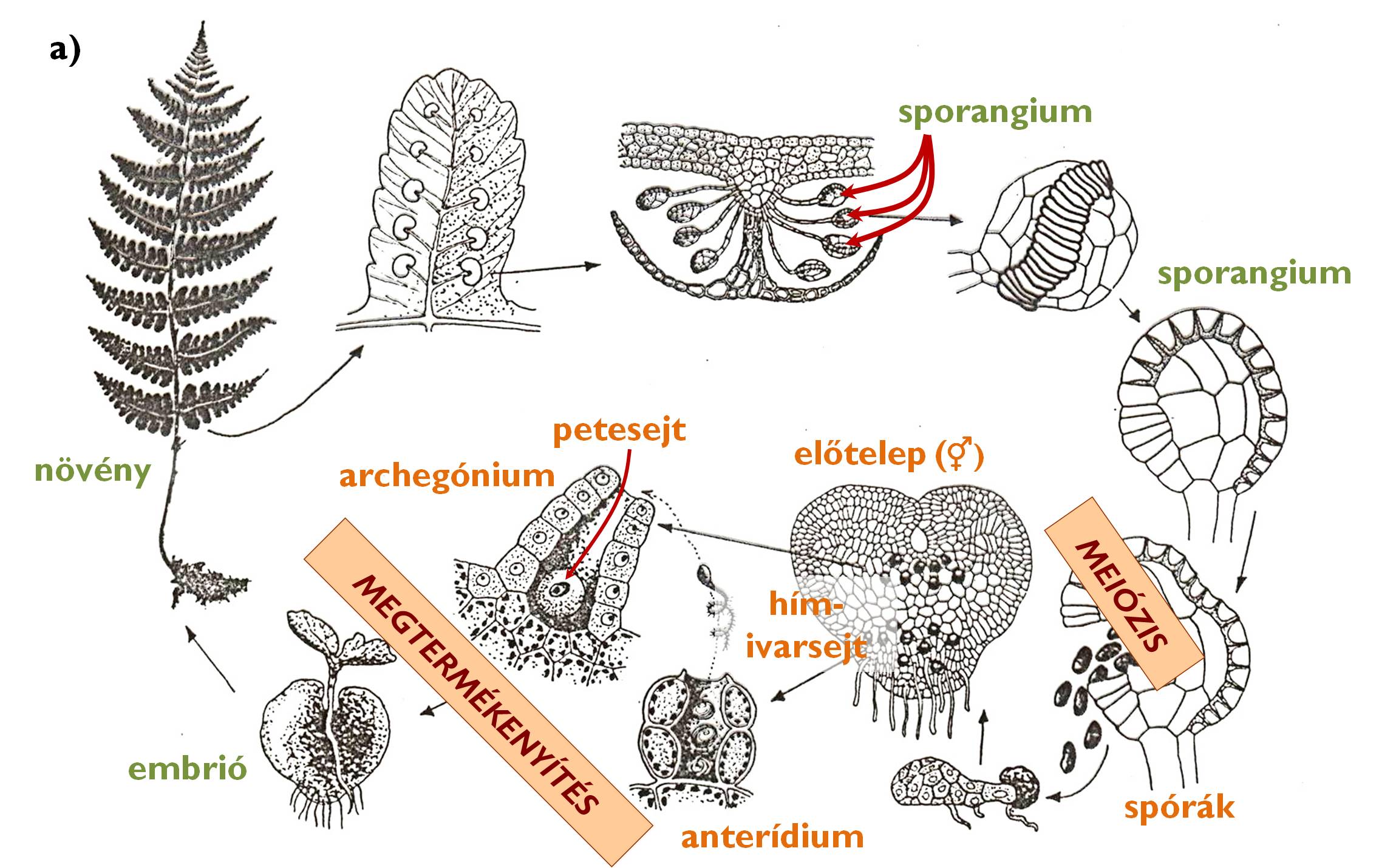
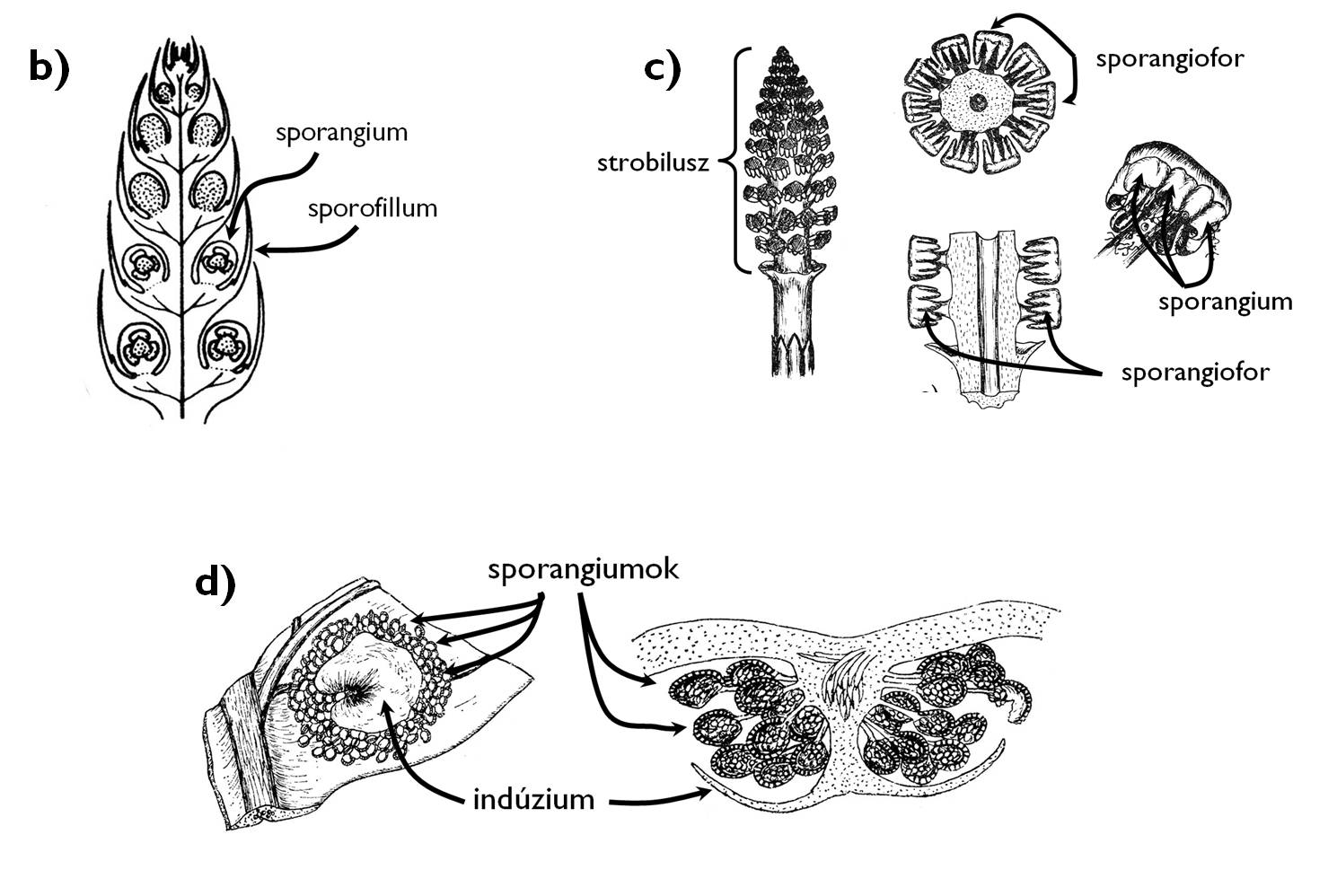
2. ábra. A harasztok életciklusának jellegzetességei.
a) a harasztok általános életciklusa a páfrányok példáján (Dryopteris filix-mas [erdei pajzsika]);
b) a korpafüvek strobilusza; c) a zsurlók strobilusza; d) a páfrányok szóruszai.
(a: Hortobágyi nyomán, b-d: Sárkány és Szalai nyomán)
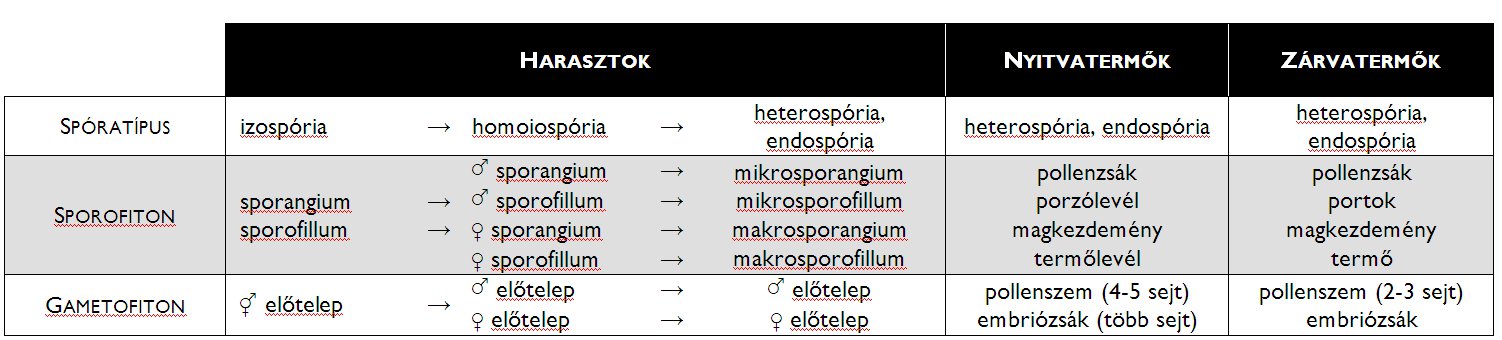
Az általános alapvonások ellenére az egyes mai harasztcsoportok eltérnek egymástól a körükben előforduló spóratípusok tekintetében, abból a szempontból, hogy sporofillumaik fotoszintetizálnak-e (sporotrofofillumok) valamint a sporofillumok elhelyezkedésében.
A Lycopodiophyta törzsbe [korpafűfélék] tartozó korpafüvek (Lycopodiales) izospórások, azaz előtelepükön anteridiumok és archegóniumok egyaránt kialakulnak. A protallium – kialakulását követően – évekig a talajban él, a vele szimbiotikus kapcsolatban élő gombafonalak táplálják. A zigótából kialakuló növényen a sporofillumok hajtásvégi füzért, ún. strobiluszt alkotva helyezkednek el, a sporangiumok pedig ezek tövében egyesével jönnek létre (2. b ábra). A csipkeharasztok rendje (Selaginellales) szintén ennek a törzsnek a tagja. Minden fajuk sporotrofofillumokkal rendelkező, heterospórás növény. A felemelkedő hajtásvégek csúcsi részén a hím jellegű spórákat termelő mikrosporangiumok, alattuk a női makrosporangiumok helyezkednek el. A csoport képviselőinek szaporodásbiológiai jellegzetessége az endospória. Ez azt jelenti, hogy a spórák kiszóródását követően a fejlődésnek induló előtelep a spórafal védelmében jön létre. A makrospórában kialakuló archegóniumok csak nyúlványaikat bocsátják ki a spórafalon kívülre, és ezen tapadnak meg a mikrospóra belsejében elhelyezkedő hím előtelepről érkező hímivarsejtek.
A Monilophyta törzs [páfrányok és zsurlók] egyik osztályát a zsurlók alkotják (Equisetopsida). Homoiospórás fajok, előtelepük kétlaki. A hím előtelepen csillókoszorús hímivarsejtek alakulnak ki. A megtermékenyítés után a női előtelepen fejlődésnek induló embrióból létrejövő zsurlónövény steril és fertilis hajtásokat egyaránt létrehozhat. A fertilis hajtás végén sporangiumtartóvá (sporangiofórum) módosult, felülnézetben hatszögletű sporofillumok alkotnak strobiluszt (2. c ábra). A sporangiofórumok fonáki (a strobilusz tengelye felé néző) oldalán hat sporangium helyezkedik el. A spóraképzés időszaka alatt a sporangiumtartók szorosan egymás mellett állnak, ám a spóraszórásra eltávolodnak egymástól, ezzel lehetővé téve a spórák terjedését. A spórák zöld színűek, faluk külső rétege felhasadva négy hosszabb, higroszkópos mozgásra képes „kart” (haptera) alkot. Párás környezetben a hapterák feltekerednek a spóra felületére, elősegítve ezzel a spórák mozgását, míg száraz körülmények között hosszan kinyúlva összeakadnak egymással, s ilyenkor a spórák nehezebben terjedhetnek.
A Monilophyta törzs többi osztályába különféle páfránycsoportok képviselőit soroljuk. Változatos felépítésük ellenére szinte mindegyikre jellemző, hogy izospóráikból kialakuló szív alakú előtelepükön anteridiumok és archegóniumok egyaránt megjelennek. A spórák barnás színű sporofillumokon (pl. Matteuccia [struccpáfrány]) és sporotrofofillumokon (pl. Nephrolepis [szobapáfrány], Dryopteris [pajzsika]) egyaránt kialakulhatnak. A sporangiumok kerekded csoportokban, szóruszokat alkotva helyezkednek el a levelek fonákán (2. d ábra). A szóruszokat – a legtöbb faj esetében – egy hártyás képlet, a fátyol (indúzium) borítja. A szóruszok és az indúzium megléte, alakja, mérete faji bélyeg a páfrányok körében. Különleges csoportot képvisel a páfrányok között a vízi páfrányokat magába foglaló Salviniales rend [rucaörömfélék]. Az ide tartozó fajok heterospórásak, ráadásul rájuk is jellemző – a csipkeharasztokhoz hasonlóan – az endospória jelensége.
A nyitvatermők szaporodása
A nyitvatermők előfutárainak (elő-nyitvatermők) evolúciós története igen korán elvált a harasztokétól, ám adaptív szétterjedésükre a földtörténeti óidő szélsőségesen száraz éghajlatú perm időszakáig kellett várni. Ezen klimatikus viszonyok között jelentettek ugyanis egyértelműen előnyt a nyitvatermőkre jellemző evolúciós újítások: a víztől függetlenné váló szaporodás és a gametofiton védelmére kialakult magkezdemény valamint a fejlődő embriót védő mag.
Az első nyitvatermők ősi heterospórás (és endospórás) harasztcsoportok leszármazottai voltak, sporofitonjukon tehát hím jellegű, mikrospórákat termelő mikrosporangiumok és női elköteleződésű spórákat termelő makrosporangiumok helyezkedtek el. Az egyik legfontosabb újítás az volt, hogy a makrosporangium körül a vegetatív szövetek (feltehetően steril telómák) összeolvadásával kétrétegű burok (integumentum) alakult ki. Ez a burokkal övezett makrosporangium a magkezdemény (3. ábra). A magkezdeményben termelődő makrospórák többé már nem hullnak ki a sporangiumból, hanem itt, a makrospóra helyén jön létre a nyitvatermők női jellegű gametofitonja is (endospória!). A néhány sejtből álló, a makrospóra falán belül kialakuló női gametofiton az embriózsák. Természetesen a magkezdemény is – hasonlóan a harasztok sporangiumaihoz – sporofillumon ül. A nyitvatermőktől kezdve ezt a makrosporofillumot termőlevélnek nevezzük (2. táblázat, 4. ábra).
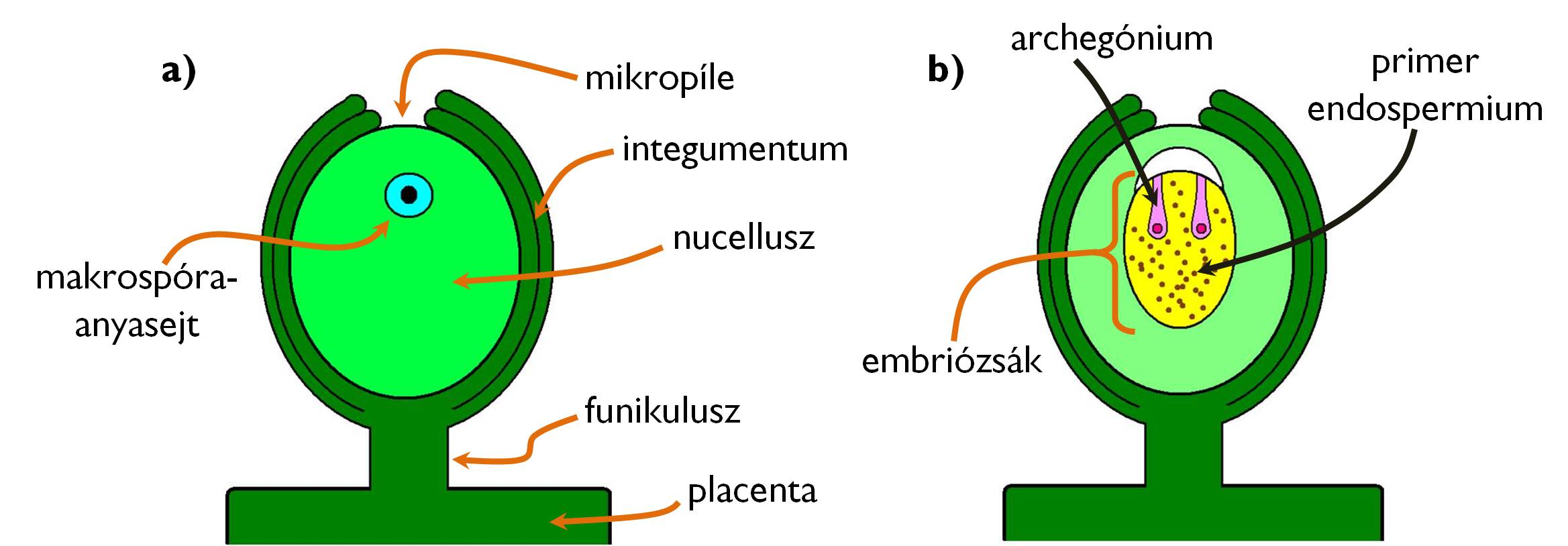
3. ábra. A nyitvatermők éretlen (a) és érett magkezdeménye (b).
A magkezdeményt a termőlevélhez a köldökzsinór (funikulusz) kapcsolja (a funikulusz csatlakozási helyét nevezzük kalazának). A termőlevél magkezdeményeket hordozó felülete a magléc (placenta). A magkezdeményt beborító integumentum köldökzsinórral ellentétes oldalán egy kis nyílást, a csírakaput (mikropile) találjuk. Ezen a nyíláson át jutnak be a hímivarsejtek a magkezdemény belsejébe. A magkezdemény fő tömegét a nucellusz („magkezdemény teste”) teszi ki. A felsorolt képletek valamennyien az anyai sporofitonnal egyező, diploid információtartalmú sejtek.
A nyitvatermők magkezdeményének speciális vonásai
A nucellusz belsejében – miután a magkezdemény elér egy adott fejlettségi állapotot – elkülönül egy kitüntetett, diploid sejt, a makrospóra-anyasejt. Ez meiózissal osztódva négy haploid makrospórát hoz létre, amelyek közül három elpusztul, és csak az egyik fejlődik később embriózsákká. Ez a makrospóra számos mitózissal osztódva egy sok haploid sejtmagból álló tömeget, a primer endospermiumot hozza létre, amiben csak később alakulnak ki a sejthatárok. A cellularizációval párhuzamosan az endospermium felületén kialakul két archegónium is, benne egy-egy petesejttel. A primer endospermium és az archegóniumok együttese (az embriózsák) képviseli a nyitvatermők női gametofitonját.
A mikrosporangium fejlődésében is történtek változások a harasztokéhoz képest. A benne kialakuló mikrospórák gametofitonná differenciálódása itt is a sporangium belsejében történik, és a spórafalon belül létrejött (endospória!), néhány sejtes gametofiton hagyja el a sporangiumot. A hím jellegű gametofiton nem más, mint a virágporszem (pollen), a mikrosporangiumot pedig már pollenzsáknak nevezzük. A mikrosporofillum neve is megváltozik, a nyitvatermőktől kezdve porzólevélről beszélünk. (A porzó- és a termőlevél az ivarlevelek.)
A nyitvatermők hím gametofitnja
A termő- ill. porzólevelek a nyitvatermők esetében is füzért (strobiluszt) alkotnak, amelyeket azonban a nyitvatermőktől kezdődően már (takarólevelek nélküli) virágnak tekintünk. E virágok minden esetben csak egyféle ivarlevélből állnak, azaz egyivarúak. A Pinophyta (fenyőalkatúak) csoportjában a virágok a legtöbb esetben tobozt alkotnak, ami a porzóleveles toboz esetében a virágnak, a termőleveles toboz esetében a virágzatnak felel meg. A termőleveles tobozvirágzat ugyanis összetett, erre utal, hogy az elfásodó vagy elhúsosodó termőlevelek (tobozpikkelyek, termőpikkelyek vagy magpikkelyek) tövében steril fellevelek (meddő pikkelyek) helyezkednek el.
A női és a hím ivarú virágok elhelyezkedhetnek egy egyeden (egylaki fajok – pl. Abies [jegenyefenyő], Larix [vörösfenyő]) vagy különböző egyedeken is (kétlaki fajok – pl. Ginkgo [ginkgófa], Juniperus [boróka]). A megporzás a nyitvatermők körében szinte kizárólag szél segítségével történik. A fenyők esetében ezt a megporzási módot segíti a légzsákos pollenszemek kialakulása: ekkor a pollenfal két rétege között levegővel telt üreg nyílik. A pollenkamrába, majd a nucellusz felületére érkező pollenszem tömlősejtje kialakítja a pollentömlőt, amelyben kezdetben a testi sejt, az embriózsákhoz közelítve pedig – a testi sejt mitózisát követően – a két hímivarsejt halad. Az archegoniális kamrába kerülő ivarsejtek csillóik segítségével jutnak el az archegóniumokig, majd a petesejtekig. A megtermékenyítés tehát víztől függetlenné válik. A megtermékenyítésre többnyire csak a megporzást követő évben kerül sor, a Ginkgo esetében a már földre hullott magkezdemények („éretlen magvak”) belsejében.
A megtermékenyítést követően megindul a magkezdemény maggá alakulása. A petesejt(ek)ből zigóta, majd embrió lesz, az integumentum rétegeiből a kétrétegű maghéj, míg a gametofiton többi sejtje táplálószövetté lesz (primer endospermium). A mag felépítésében tehát különböző genetikai információtartalmú sejtek vesznek részt: anyai sporofitonnak megfelelő, diploid sejtek (maghéj), a gametofiton haploid sejtjei (primer endospermium) és az utód sporofiton diploid sejtjei (embrió). A mag kialakulása után – hormonális (elsősorban abszcizinsav) hatásra – beáll a magnyugalmi állapot, ami később a csírázás megindulásával szakad meg.
A zárvatermők szaporodásának alapvonásai
A magvaspáfrányok evolúciós története a nyitvatermők felé vezető ág mellett egy azzal párhuzamos vonalon elvezetett az első zárvatermők kialakulásához. E növények esetében a makrosporangium és az embrió védelme a nyitvatermőkénél is fokozottabbá vált: a magkezdemények nem nyitott termőleveleken ülnek, hanem a sporofillum szöveti állománya körbezárul a sporangium körül. A magkezdeményt magába záró termőlevélből így kialakul a termő, míg a pollenzsák körül záródó porzólevélből a portokot hordozó porzó (2. táblázat, 4. ábra).
Ezzel párhuzamosan a gametofiton szakasz további redukciója figyelhető meg. A hím vonalon immár csak két sejt (vegetatív és generatív sejt) képviseli a haploid szakaszt, míg a női vonalon az embriózsák sejtjeinek száma csökken jelentősen. A zárvatermőknél az archegónium is eltűnik! Ugyanakkor a zárvatermők virágjában kialakulnak a takarólevelek, ami lehetővé teszi a rovarmegporzás létrejöttét.
Evolúciós újítás a kettős megtermékenyítés folyamata is. Habár a nyitvatermők mikrospórájának osztódásaival is két hímivarsejt jött létre, ott ezek az embriózsák két petesejtjét termékenyítették meg. A zárvatermők esetében a két hímivarsejt egyazon embriózsák két eltérő jellegű sejtjével egyesül. A zigóta mellett a mag táplálószövete (szekunder endospermium) is a két szülő génállományát hordozó sejtekből áll.
A termőlevelek záródásával természetesen nemcsak a magkezdeményt veszik körül további anyai (diploid) szövetrétegek, hanem azok a kialakuló magvak körül is megmaradnak. Így jön létre a megtermékenyítést követően a magvak védelmére és terjesztésére szolgáló termés.
A zárvatermők ivaros folyamatait részletesen a fejezet további részeiben tárgyaljuk.
2. táblázat. A szaporodás evolúciója.
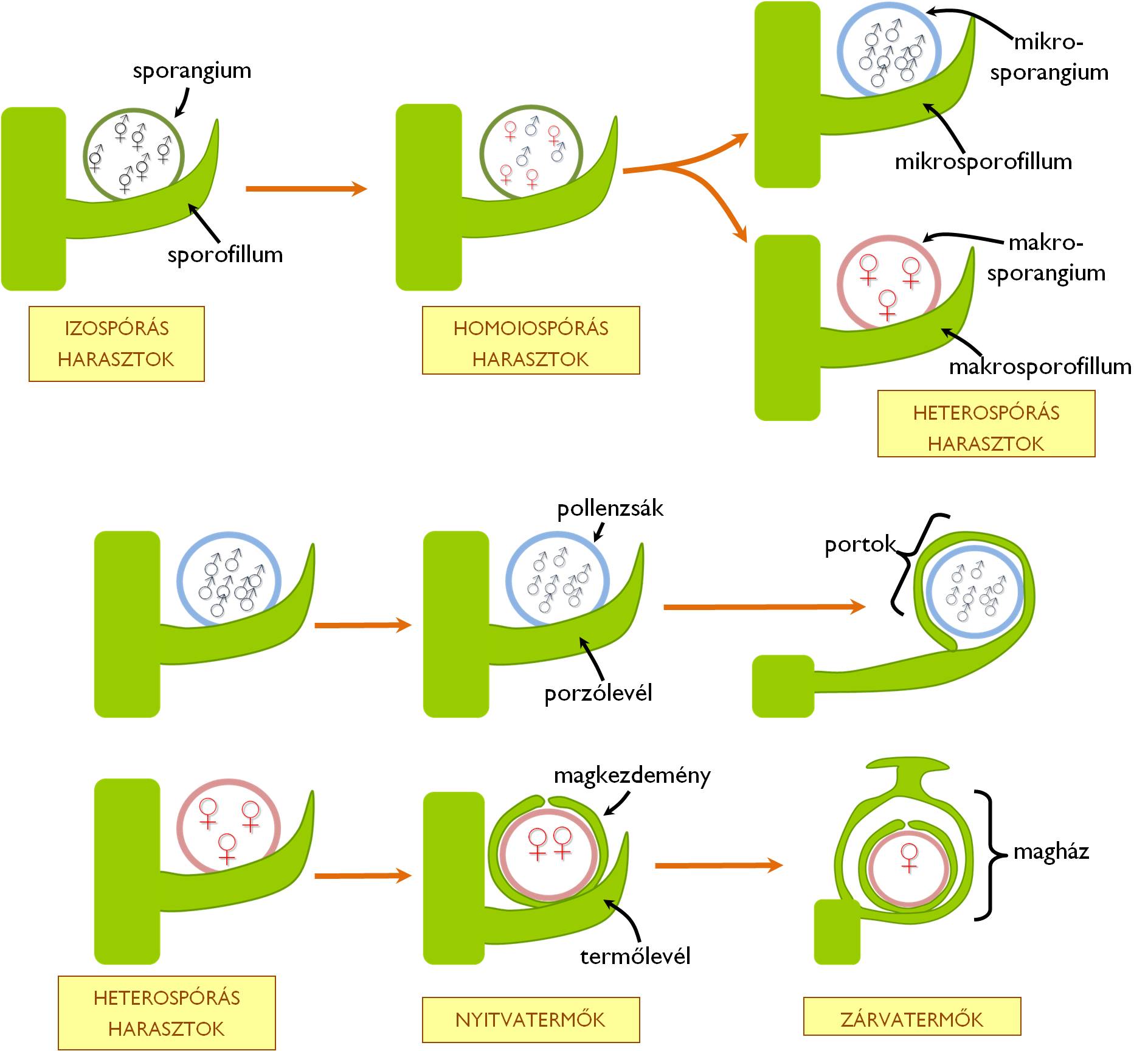
4. ábra : Animáció a növényi szaporítószervek leszármazási kapcsolatairól. Az animáció során a fenti ábrán szereplő képletek egymásba alakulása látszik, megjelenő feliratokkal.
Az ivartalan szaporodás és szaporítás módjai
Az ivartalan szaporodás és ivartalan szaporítás közös célja a lehető legnagyobb utódszám elérése a természetes vagy mesterséges (emberi) szelekció nyomán kialakult, előnyös génállomány megőrzése mellett. A kertészeti gyakorlat felhasználja az ivartalan szaporodás természetes szerveit is, például a vegetatív szaporodásra szolgáló szervmódosulatokat: a fiókhagymákat (sarjhagymákat) (Allium sativum [fokhagyma]), sarjgumókat (Lilium bulbiferum [tűzliliom]), elevenszülő leveleken (Kalanchoe tubiflora [ernyős sarjika], K. daigremontiana [mexikói sarjika]), indákon (Fragaria × ananassa [szamóca], Sempervivum tectorum [házi kövirózsa]), tarackokon ( Convallaria majalis [gyöngyvirág]) vagy legyökerező heverő szárak szárcsomóin (Vinca major [nagy télizöld]) megjelenő sarjnövényeket, a gyökér- vagy tősarjakat (pl. Salix [fűz], Populus [nyár] fajok), leválasztott gumókat (Solanum tuberosum [burgonya]) és feldarabolt rizómákat (Iris [nőszirom] fajok). Emellett azonban speciális, mesterséges szaporításmódok is ismeretesek (5. ábra).
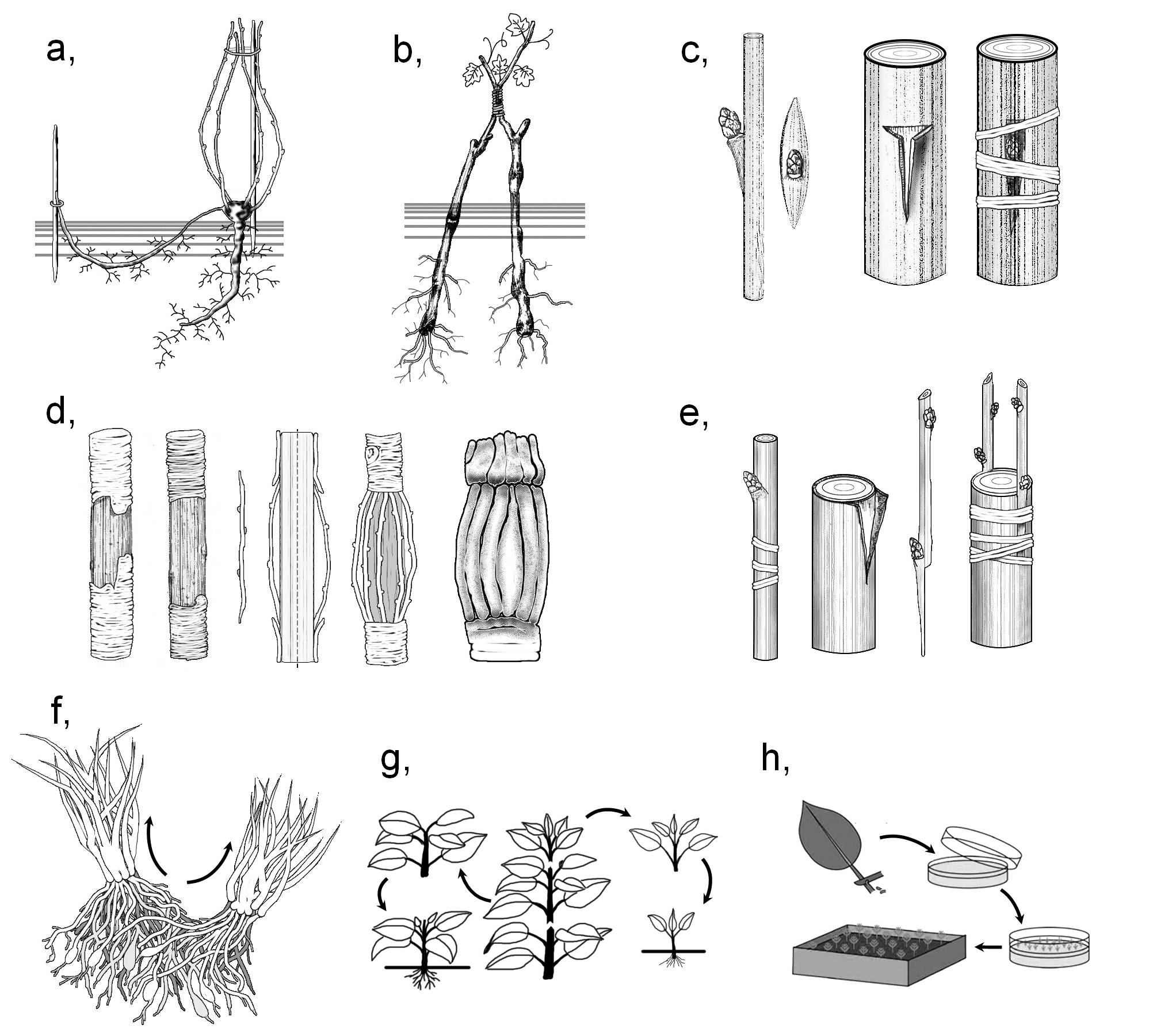
5. ábra. Az ivartalan szaporítás lehetőségei.
a) bujtás; b) ablaktálás; c) szemzés; d) áthidalás; e) oltás; f) tőosztás; g) dugványozás; h) klónozás.
(a-e: Kárpáti nyomán, f-h: internetes források alapján; f - http://answers.com, g - http://http://pubs.ext.vt.edu, h - http://bbc.co.uk)
A bokrosan növekedő, sarjtelepet képező, többnyire évelő vagy cserjés növények nagyobb egységekre vagy önálló, gyökeres hajtásokra történő szétválasztása a tőosztás. Tőosztásnak tekintjük a tő- és gyökérsarjak leválasztásával valamint a föld alatti szárak feldarabolásával történő szaporítást is. A bujtás esetében az anyatő valamely föld feletti hajtását nedves talajba hajlítjuk és megvárjuk, míg a nedves környezetbe került szárcsomókon járulékos gyökerek fejlődnek, rügyei pedig hajtásokká alakulnak. Ekkor a klónok már leválaszthatók az anyanövényről. (Bujtással főleg fás szárú növények szaporíthatók.)
A dugványozás valamely vegetatív szerv levágását, majd vízben vagy nedves gyökereztető közegben (tőzeg, homok, perlit) történő meggyökereztetését jelenti. A sebzési felület mentén ekkor többnyire sebkallusz jelenik meg, majd ebből fejlődnek ki a járulékos gyökerek. A hajtás regenerációja már meglévő vagy járulékos rügyekből történik. A felhasznált szerv alapján megkülönböztetünk szárdugványt (ezen belül fás vagy félfás illetve (zöld)hajtás dugványt), levéldugványt (pl. Begonia [begónia], Peperomia [peperómia], Sansevieria [anyósnyelv]), levélnyéldugványt (pl. Saintpaulia [afrikai ibolya]), gyökérdugványt (pl. Armoracia lapathifolia [torma], Papaver orientale [keleti mák]).
A szervátültetés (transzplantálás) során egy élő növényről (nemes) leválasztott szervet egy másik növényen (alany) ejtett sebbe helyezünk. Ezzel elérhetjük, hogy a kívánt kertészeti jellemzővel (megfelelő termésméret, beltartalom, virágszín stb.) rendelkező, de többnyire érzékenyebb nemes az ellenállóbb alanynak köszönhetően szélsőségesebb körülmények között is termeszthető legyen. Ezen a módon biztosítható a nemes gyors és tömeges elszaporítása is.
Az átültetést követően a sebzés hatására a két növény szerveiben – a kezdeti, felszíni részeket érintő sejtelhalást követően – sebkambium jön létre másodlagosan, amelynek osztódása egy parenchimatikus jellegű köztes (intermedier) szövetet hoz létre. Ez részben áttöri, részben egymásnak feszíti a sebzési felszíneket, így egy szabad szemmel is látható elválasztó réteg alakul ki. Az intermedier szövet később ezt a réteget is áttöri, miközben sejtjei osztódóképessé válnak, majd differenciálódni kezdenek. Eközben a seb külvilág felé eső részein záró funkciójú sebpara képződik. A folyamat utolsó fázisában az intermedier szöveten keresztülhúzódó parenchimahidak jönnek létre az alany és a nemes között, ezzel a két növény élő sejtjeinek kapcsolódása is megtörténik. Eközben lassan megkezdődik a két partner kambiumzónájának beterjeszkedése a köztes szövetbe. Az alany és az oltvány kambiuma végül összeér, folytonossá válik, és ekkortól már közös osztódószöveti rétegként gyarapítja tovább az oltvány mindkét részének szöveti állományát. Ezzel a két növényi szerv végképp összeforr.
A transzplantációnak a kertészeti gyakorlatban többféle módja ismeretes. A szemzés során egyetlen rügy, oltás esetében egy hajtásrészlet átültetésére kerül sor. Az ablaktálás alkalmával a nemes hajtása összefüggésben marad az anyanövénnyel. Ekkor a két növényen hasonló méretű sebet ejtünk, amelyeket pontosan összeillesztünk, és erősen összekötözzük azokat. A nemes hajtásrészt csak a szövetek folytonossá válása után választjuk le az anyáról. Ezzel a módszerrel olyan érzékenyebb növényi szervek transzplantálására is van mód, amelyek a szöveti folytonosság kialakulása előtt elpusztulnának. Az áthidalás sérült növényi részek (pl. korhadt ágak) pótlására szolgál. Ekkor az elhalt részt átívelő ágakat a sérülés mindkét oldalán az alany hajtástengelyére oltjuk.
A vegetatív szaporítás legújabb módszere a klónozás, amely amellett, hogy lehetővé teszi a korábban bemutatott módszerekkel nehezen szaporítható növények (pl. orchideák vagy rovarfogó növények) propagálását, lehetőséget nyújt azok génállományának megváltoztatására is. Ebben az esetben az anyanövényről izolált (többnyire alapszöveti vagy osztódó szöveti) sejteket mesterséges (in vitro) körülmények között, hormontartalmú táptalajon tartjuk életben. Kezdetben egy differenciálatlan sejtekből álló kallusz jön létre, amelyből azonban a táptalajhoz adott hormonok (elsősorban auxinok és citokininek) arányának megfelelő megválasztásával regeneráltathatók a növény vegetatív szervei. Természetesen egyetlen anyanövény sejtjeiből több kallusz is előállítható, ráadásul a kalluszok feldarabolására is van lehetőség, így a módszer ténylegesen alkalmas a szaporításra. (A kalluszt alkotó sejtek génállománya megfelelő molekuláris genetikai módszerekkel változtatható meg, ezek bemutatása azonban nem ennek a jegyzetnek a feladata.)
Az ivaros szaporodás
A virág
A virág fogalma és evolúciója
A virág ivaros szaporodásra szolgáló, módosult leveleket hordozó, korlátolt növekedésű, rövid szártagú hajtásrészlet. Az evolúció során a nyitvatermőknél jelenik meg – azaz kialakulása a magkezdemény és a pollenzsák létrejöttével állítható párhuzamba. (Megjegyzendő mindazonáltal, hogy egyes kutatók csak a zárvatermők hasonló felépítésű szaporítószerveit nevezik virágnak.)
A virág evolúciós eredete: a pszeudanthium és az euanthium elmélet
A virág felépítése
A zárvatermők virága a módosult hajtástengelyből (virágtengely) és az ezen elhelyezkedő, a szaporodást szolgáló módosult levelekből ill. sporofillumokból áll. A virágtengely részei a virágot hordozó szárrész, a kocsány, valamint ennek kiszélesedő, a leveleket hordozó szakasza, a vacok. A viráglevelek külső köreit a takarólevelek (csésze- és szirom- vagy lepellevelek) alkotják, míg a sporofillumok evolúciós leszármazottai az ivarlevelekből kialakult porzó illetve termő.
Azt a virágot, amelyben valamennyi alkotórész (takaró- és ivarlevél) megtalálható, teljes virágnak nevezzük. Ellenkező esetben a virágot hiányos virágnak hívjuk. A csupasz virágnak a takarólevelei hiányoznak (pl. Salix [fűz], Populus [nyár]). A meddő virágnak ivarlevelei nincsenek, funkciója többnyire a rovarcsalogatás. Az egyféle ivarlevelet tartalmazó virág az egyivarú virág. Ha a virág porzót és termőt egyaránt tartalmaz, kétivarúnak mondjuk. Az egyivarú virág, attól függően, hogy melyik ivarlevél található meg benne, lehet porzós vagy termős virág. Néhány növényfaj (pl. Fraxinus angustifolia subsp. danubialis [magyar kőris]) virágaiban annak ellenére, hogy mindkét ivarjelleget hordozzák, a termő vagy a porzó redukálódik. Ekkor funkcionálisan egyivarú virágról beszélünk. Más növénynemzetségek egyedein (pl. Acer [juhar], Vitis [szőlő], Morus [eperfa], Sorghum [cirok]) egy- és kétivarú virágokat egyaránt találunk; a jelenség neve felemás-virágúság.
Az egyivarú virágot hordozó növények lehetnek egylakiak, ha a termős és porzós virágok egyazon egyeden megtalálhatók (pl. Fagaceae [bükkfafélék], Betulaceae [nyírfafélék], Cucurbitaceae [tökfélék], Zea mays [kukorica], Begonia sp. [begónia]). A kétlaki növények termős és porzós virágai külön egyedeken helyezkednek el (pl. Salicaceae [fűzfafélék], Urtica dioica [nagy csalán], Humulus lupulus [komló], Cannabis sativa [kender]).
A virág kialakulása és a virágnyílás
A virágos növényeket első virágaik megjelenésétől kezdve tekintjük ivarérettnek. Az egyéves növények esetében ez a csírázással azonos évben következik be, míg többéves fajok esetében a virágzást megelőző évben alakulnak ki a virágot tartalmazó rügyek, a generatív hajtáscsúcsot tartalmazó virág- vagy vegyesrügyek.
A generatív hajtáscsúcs kialakulása
Hányszor hoz virágot egy növény?
A virágnyílás időpontja változó lehet mind a tenyészidőszak folyamán, mind egyetlen napon belül is. Fás szárú növényeink virágozhatnak lombfakadás előtt (pl. Cornus mas [húsos som], Prunus amygdalus [mandula]) azzal egy időben (pl. Acer platanoides [korai juhar], Prunus persica [őszibarack]), vagy lombfakadás után (pl. Cornus sanguinea [veresgyűrű som], Vitis vinifera [kerti szőlő]). Ezt természetesen megszabja a generatív részeket tartalmazó rügyek minősége is. A lombfakadás előtt virágzó fajoknak mindig vannak csak virágkezdeményeket tartalmazó virág(zati) rügyeik is, míg a levél- és virágkezdeményeket egyaránt tartalmazó vegyes rügyekkel rendelkező fajok csak lombfakadással egy időben, vagy azt követően bonthatják virágaikat. (Megjegyzendő, hogy a levél- és virágrüggyel egyaránt rendelkező fajok esetében is bomolhat egyidőben a kétféle rügytípus, ami lombfakadással egyidejű virágzást jelent – pl.: Prunus persica [őszibarack].)
A virágzás időpontját a megvilágított órák száma is befolyásolhatja. A trópusi, szubtrópusi területekről származó rövidnappalos növények (pl. Chrysanthemum × hortorum [krizantém], Coffea arabica [kávé], Oryza sativa [rizs]) virágzásához nagyjából 12 órás megvilágítás és legalább 12 órán át tartó folyamatos sötétség szükséges (3. táblázat). E fajok nálunk nyár derekán vagy annál is később virágoznak. Ezzel szemben a hosszúnappalos növények, amelyek a mérsékelt ill. a hideg övből származnak, hazánkban tavasszal vagy nyár elején bontják virágaikat, mivel virágzásukhoz 12-14 órán át tartó megvilágítást igényelnek (pl. Pisum sativum [borsó], Triticum aestivum [búza], Lactuca sativa [saláta]). Ezen fajok számára a nappalok megnyúlása jelzi a kedvezőtlen, hideg időszak végét, a csapadékos vegetációs időszak megindulását. (+info) (A megvilágítás hosszától függetlenül virágzó fajokat napneutrális növényeknek nevezzük.)
3. táblázat. A rövid- és hosszúnappalos növények összehasonlítása.
|
Rövidnappalos növény |
Hosszúnappalos növény |
|
|
Származás |
Trópusok, szubtrópusok (mediterrán) |
Mérsékelt és hideg öv |
|
Virágzása hazánkban |
Nyár végén, ősszel |
Tavasszal, nyár elején |
|
A virágzás feltétele |
min. 12 óra folyamatos sötétség |
8-10 (max. 12) órán át tartó sötétség |
|
Példafajok |
Chrysanthemum × hortorum [krizantém] Coffea arabica [kávé] Nicotiana tabacum [dohány] Gossypium hirsutum [gyapot] Oryza sativa [rizs] Saccharum officinarum [cukornád] Euphorbia pulcherrima [mikulásvirág] Kalanchoe blossfeldiana [korallvirág] |
Dianthus caryophyllus [kerti szegfű] Hyoscyamus niger [beléndek] Secale cereale [rozs] Lolium perenne [angolperje] Trifolium pratense [réti here] Campanula sp. [harangvirág] Pisum sativum [borsó] Hordeum vulgare [árpa] Lactuca sativa [saláta] Triticum aestivum [búza] |
Mikor nyitja virágait a növény?
A virágrészek szövettana
A virág alkotórészeinek anatómiája egyértelműen mutatja, hogy valójában egy módosult hajtás szervei alkotják. A virágkocsány szöveti felépítése a hajtástengely egyéb részeinek megfelelő, azzal a különbséggel, hogy az esetek többségében a virágkocsányban már fokozódik a szilárdító szövet mennyisége (szklerenchimatikus nyalábhüvely, hipodermisz, kollenchima-rétegek). Ezek mennyisége a terméskötéssel tovább nő, annak érdekében, hogy a tengely képes legyen megtartani a termése tömegét.
A takarólevelek felépítésének alapszabása a lomblevelekéhez hasonló (6/a ábra): a két epidermisz között parenchimatikus jellegű alapszövetből álló mezofillumot, benne kollaterális zárt szállítónyalábokat találunk. A csészelevelek (illetve egyes esetekben a külső elhelyezkedésű, zöld színű lepellevelek) anatómiája emlékeztet leginkább a lomblevelekére: bőrszövetükben működő gázcserenyílások, esetenként szőrök helyezkednek el, mezofillumukban klorenchima fordul elő, néha a paliszád réteg is felismerhető (bár gyakoribb a homogén izolaterális felépítés). A sziromlevelek és a lepellevelek epidermiszében működő sztómák már nem fordulnak elő, viszont gyakori a papillák kialakulása felszínükön. A mezofillum állományát izodiametrikus, vékony falú parenchimasejtek alkotják, bennük a szirom színétől függően különféle színanyagok (lilás-pirosas antociánok, sárga betalainok vagy flavonoidok, narancsszínű vagy piros karotinoidok, zöld klorofill) halmozódhatnak fel. (A fehér szín esetében a takarólevél valamennyi fénykomponenst visszaveri; ennek külön színanyaga nincsen.) A szállítónyalábok a takarólevelekben erősen redukálódnak, többnyire csak néhány szállítóelemet tartalmaznak.
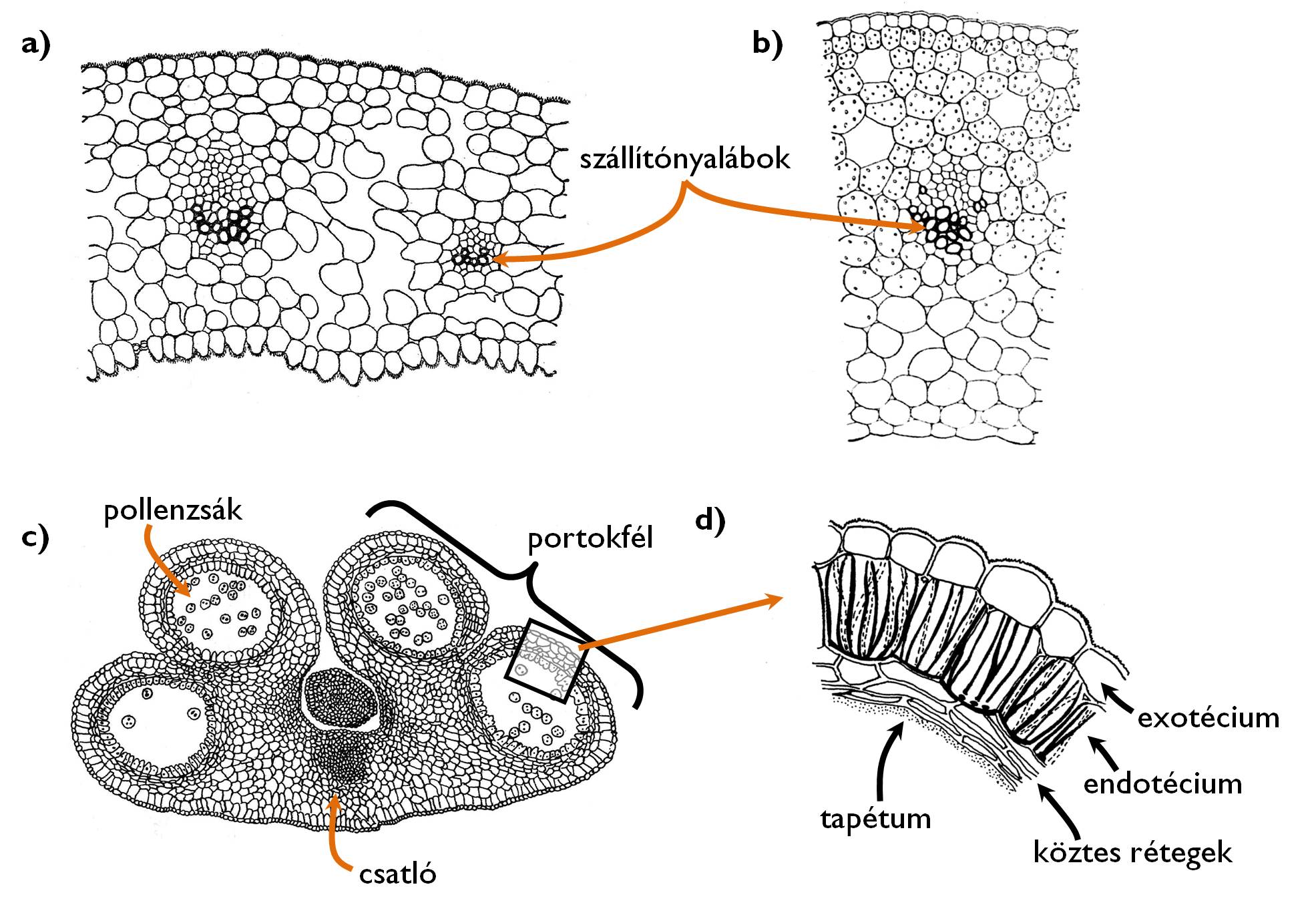
6. ábra. A virágrészek szövettana.
a) a csészelevél felépítése (Helleborus [hunyor]); b) a magház falát alkotó termőlevél felépítése (Tulipa [tulipán]); c) a portok szerkezete (Lilium [liliom]); d) a portokfal rétegei.
(Sárkány és Szalai nyomán)
A porzótájat (andröceum) a porzólevelekből kialakult porzók (sztamen) alkotják. Ezek két részből, a porzószálból (filamentum) és a portokból (antera) állnak (6/c-d ábra). A porzószál a két portokfelet (téka) összekötő csatlóba (konnektívum) torkollik bele, egyetlen kollaterális szállítónyalábja ennek belsejében folytatódik. Egy portokfélben két pollenzsák (lokulamentum) található, egyetlen porzólevél tehát összesen négy mikrosporangiumot zár körül. A portok fala többnyire négy rétegből áll. A legkülső exotécium nem más, mint a porzólevél egyrétegű epidermisze, amit a jellegzetesen hálózatosan vastagodott falú sejtek rétege, az endotécium követ. E rétegben a pollenszemek megérése után, a szövetek vízvesztése miatt feszítő erő keletkezik, ami felrepeszti a portokokat. Az endotéciumon belül egy (néha hiányzó) átmeneti vagy köztes réteg következik, belülről pedig a tapétum béleli a lokulamentumot. A tapétum sejtjei hozzák létre a centrifugálisan vastagodó pollenfal anyagait, amelyek a vegetatív sejt falára rakódva létrehozzák a virágporszemek jellegzetes felszíni mintázatát.
A termőlevelek (karpellum) összenövésével létrejövő termő (pisztillum) képviseli a zárvatermők termőtáját (ginöceum). A termő részei a pollenszemek megtapadási helyéül szolgáló bibe (stigma), a bibét a virágból kiemelő bibeszál (sztilusz) valamint a magkezdeményeket tartalmazó magház (ovarium). A magházban levő magtanyán vagy maglécen ( placenta) ülnek a magkezdemények (ovulum).
A bibe felületét papillás mirigyszövet alkotja, ami a kiválasztott cukros váladék segítségével segíti a pollenszemek megtapadását. (+info) A bibeszál belső állományában, parenchimába ágyazva találjuk a pollentömlő előrehaladását segítő vezető szövetet. Ennek sejtjei vezetik a pollentömlőt a bibeszálat alkotó sejtek sejtfalának poliszacharid jellegű részein keresztül a magház felé. A vezető szövet sejtjei emellett a pollentömlőt is táplálják.
A placentáció és a magkezdemény változatossága
A zárvatermők mikrosporo- és mikrogametogenezise
A zárvatermők mikrosporogenezise a pollenzsákban levő sporogén szövetben létrejövő diploid mikrospóra-anyasejtek (pollen-anyasejtek) meiózisával indul (7. ábra). Az osztódás eredményeképpen kialakuló négy haploid mikrospórát egy közös kallózból álló sejtfal veszi körül, ezért azok négyesével együtt maradva tetrádokat alkotnak. (Ritkán az is előfordul, hogy az érett pollenszemek is négyesével együtt maradnak – pl. az Ericaceae családban [hangafélék].) A környezetüktől elszigetelt tetrádokat a pollenzsákot bélelő tapétum sejtjei táplálják. A kallóz sejtfal alatt már mikrospóra állapotban megindul a későbbi pollen sejtfalának szintézise.
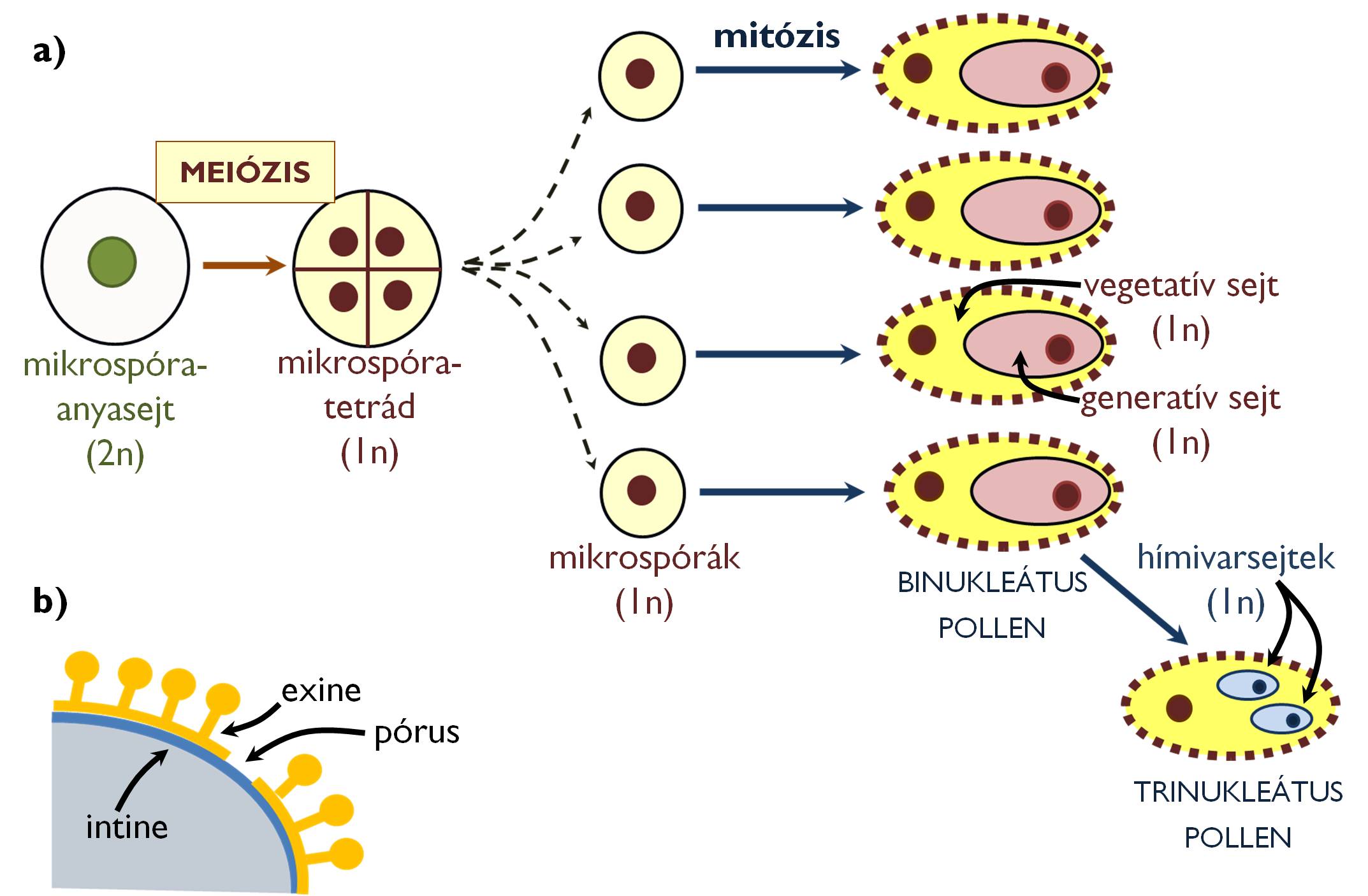
7. ábra. A zárvatermők mikrosporo- és mikrogametogenezise (a);
a pollenfal felépítése (b).
A rendkívül ellenálló, sporopolleninből álló sejtfal kialakulása közben a tetrádokat összetartó kallóz elbomlik, a mikrospórák szétesnek, sejtjeik pedig mitózissal osztódnak, létrehozva a zárvatermők hím gametofitonját képviselő két haploid sejtet: a nagyobb vegetatív sejtet, valamint az ennek belsejében elhelyezkedő, kisebb méretű generatív sejtet. A pollenszemek nagy része ebben, a kétmagvas állapotban szóródik a portokból (binukleátus pollen), míg a növényfajok mintegy harmadánál (pl. a Poaceae családban [pázsitfűfélék]) a generatív sejt már a portokban mitotikusan kettéosztódik két hímivarsejtté. (Ekkor a pollen trinukleátus, azaz három sejtmagot figyelhetünk meg benne.) A binukleátus pollen esetében a generatív sejtből csak a pollentömlőben alakulnak ki a hímivarsejtek (a mikrogametogenezis folyamata ekkor tehát a termőben fejeződik be).
A zárvatermők makrosporo- és makrogametogenezise
A zárvatermők női jellegű spórái a makrosporangiumból származó magkezdemény belsejében alakulnak ki. A magkezdemény nucelluszának mikropile felé eső végén az egyik diploid sejt makrospóra-anyasejtté (embriózsák-anyasejt) válik: mérete megnő, kallózból álló sejtfallal különül el a környező nucelláris sejtektől. A makrospóra-anyasejt meiózissal osztódva négy haploid makrospórát hoz létre.
A zárvatermők női gametofitonja több különböző módon kialakulhat, de a fajok túlnyomó többségére az ún. Polygonum-típusú embriózsák a jellemző (8. ábra). Ebben az esetben az embriózsák kialakításában csak egy makrospóra vesz részt (monospórás embriózsák), a másik három hamarosan elpusztul. A makrospóra három egymást követő mitózissal összesen nyolc haploid sejtmagot hoz létre, amelyek körül a sejthatárok csak később alakulnak ki. Az embriózsákot összesen hét sejt alkotja: a mikropile felé eső részen találjuk a petesejt apparátust a petesejttel, mellette a két segítő sejttel (szinergida), míg az ezzel ellentétes (kalazális) póluson három ellenlábas (antipod) sejt helyezkedik el; az embriózsák legnagyobb sejtje a közbülső régiót kitöltő központi sejt, amelynek magja két haploid mag összeolvadásával jön létre (tehát a többi sejttel szemben ez diploid!).
Az embriózsák kialakulásának pillanatától beszélünk érett magkezdeményről. Mivel ez a megtermékenyítésig a magház belsejében marad, a női gametofiton végig az anyanövényen, a termőben marad.
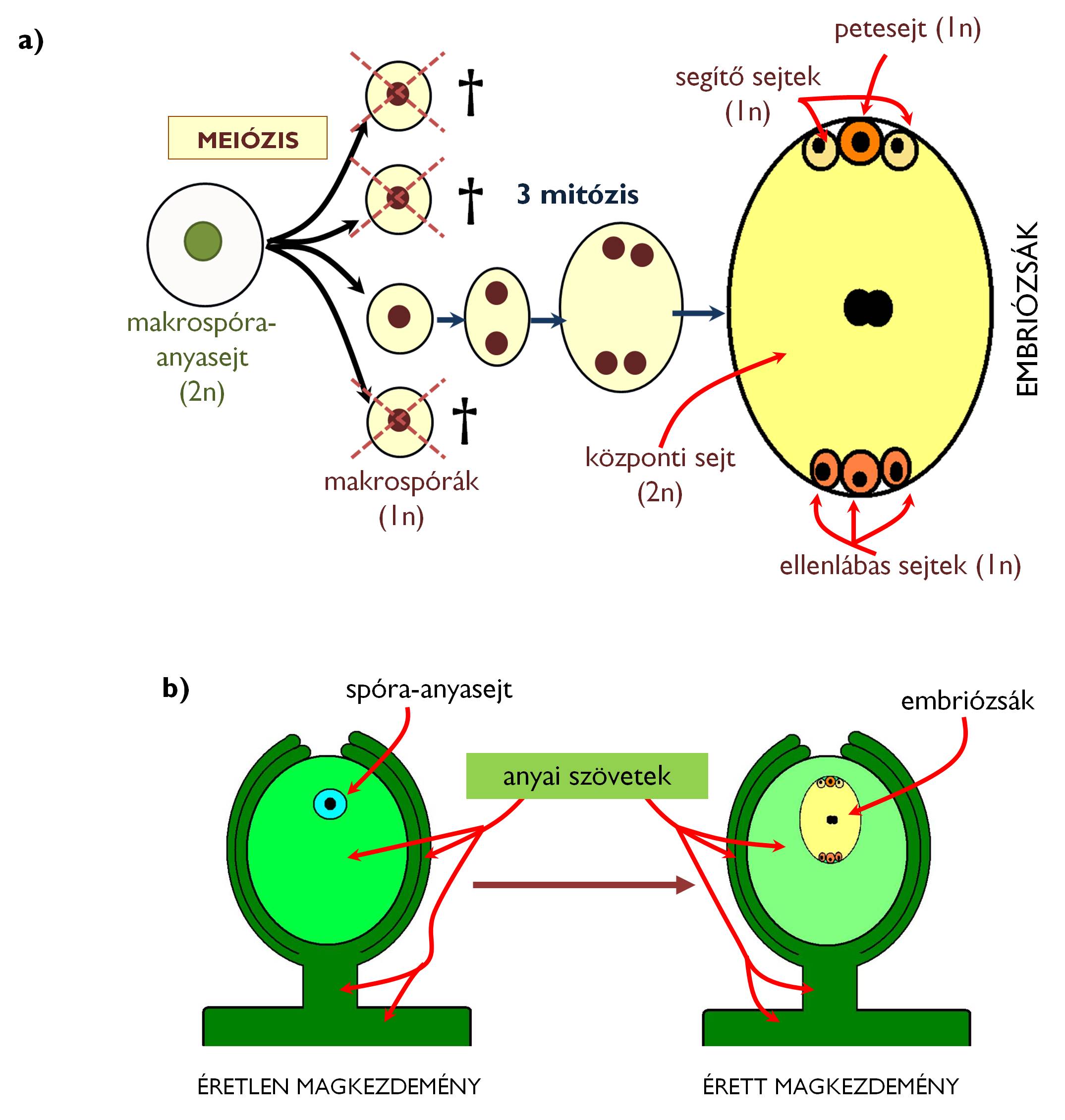
8. ábra. A zárvatermők makrosporo- és makrogametogenezise (a);
az éretlen és az érett magkezdemény (b).
A megporzás
A zárvatermők megporzása során a virágpor a termő bibéjére kerül. A virágok – a genetikai változatosság biztosítása érdekében – igen gyakran önsterilek (autosterilek), azaz a saját virágporból származó ivarsejtek nem képesek megtermékenyíteni a magkezdemény petesejtjét. Az autosterilitásnak meghatározott genetikai háttere van, és több különböző szintje lehetséges. (Például van olyan eset, hogy már a pollen nem is képes kicsírázni a bibe felületén, vagy a bibe szövetei „utasítják el” a fejlődő pollentömlőt, ami elpusztul benne.)
Az önmegtermékenyítés elkerülésének módjai
A virágporszemek csak ritkán jutnak közvetlenül a bibére. Jóval gyakoribb, hogy valamilyen külső tényező (szél, víz, állat) juttatja el azokat a termő csúcsára. A megporzás módja az ökológiai környezethez történő alkalmazkodást tükrözi, és egyértelmű morfológiai és anatómiai alkalmazkodást igényel a növény részéről.
Az önmegporzó (autogám) fajok esetében a virágok gyakran ki sem nyílnak (kleisztogámia), és a pollenszemek már a bibével érintkező portokban kicsíráznak (pl. Viola [ibolya]). Ekkor a pollentömlő áttöri a portok falát, s így jut el a bibéhez. A chazmogám-autogám fajok esetében (ahol kinyílik a virág a megporzás idejére) egyéb módokon juttatják a pollent a bibére. A fészekvirágzatúak (Asteraceae) esetében például a fejlődő bibe a portokok összenövésével létrejövő portokcsövön (szinandrium) nő keresztül mielőtt kiterülne. Mivel ezek a fajok proterandrikusak, a portokcsövön áthaladó bibe fonákára ekkor pollenszemek tapadnak. Önmegporzás ekkor még nem történik, mivel a növekedés végeztével a kétágú bibe szétterül, és a szabaddá váló bibefelszínen ekkor még nincsenek rajta a saját virágporszemek. Ha azonban nem kerül sor idegen-megporzásra, a bibekaréjok fonnyadni kezdenek, összesodródnak és egymásra tekerednek. Ekkor a bibe fonákáról eljutnak a virágporszemek a pollenkötő felületre, és megtörténik az önmegporzás.
A szélporozta (anemogám) növények portokjaiból a pollenszemek a levegőbe kerülnek és a légáramlatok juttatják el azokat a bibére. Igen sok szélporozta faj virágai lombfakadás előtt nyílnak (pl. Corylus [mogyoró], Acer negundo [zöld juhar]), hogy a lombozat ne akadályozza a pollenszemek terjedését. A lombfakadással egyidőben vagy azt követően virágzó szélporozta növények virágai az ágak végén, a levelek közül kiemelkedve helyezkednek el (pl. Populus [nyár], Fagus [bükk], Carpinus [gyertyán], Betula [nyír] nemzetségek). A virágtakaró a szélporozta fajok esetében jelentéktelen, redukált (pl. Poaceae [pázsitfűfélék], Amaranthaceae [disznóparéjfélék]) vagy gyakran teljesen hiányzik (csupasz virágok – pl. Salix [fűz]). A porzószálak, vagy a porzós virágok kocsányai igen hosszúak, mozgékonyak, a portokokban nagy mennyiségű, apró, könnyű pollenszem termelődik. A fenyőfélék (Pinaceae) esetében a virágpor terjedését a pollen sejtfalának rétegei között nyíló légzsákok is segítik a fajsúly csökkentése révén. A szélporozta zárvatermő fajok bibéje igen nagy felületű, tollas vagy ecsetszerű, hogy nagyobb eséllyel csapdába ejthesse a virágporszemeket.
A növényvilág legváltozatosabb megporzási módja a zoogámia, amikor állatok segítségével jutnak el a virágporszemek a termőre. A megporzást többnyire rovarok végzik (entomogámia), de madarak (ornitogámia), csigák (malakogámia), denevérek (chiropterogámia) vagy egyéb kisemlősök is lehetnek a megporzók (pollinátorok). A rovarok számára a virágok elsősorban táplálékot nyújtanak, a virágban vagy annak közelében elhelyezkedő nektáriumok cukros váladéka (pl. Rosaceae [rózsafélék], Lamiaceae [ajakosok]), esetleg a feleslegben termelt, fehérjében gazdag virágporszemek (pl. Papaveraceae [mákfélék], Paeoniaceae [pünkösdirózsa-félék]) formájában. Megporzóikat ezek a fajok feltűnő, színes virágtakaróikkal és ozmofóráik előállította illatanyagaikkal csalogatják magukhoz. Abban az esetben, ha a zoogám virágok aprók, azok virágutánzó virágzatba (pszeudantium) tömörülnek (pl. az Asteraceae [fészekvirágzatúak] családjának tagjai, Cornus florida [virágos som], Scabiosa ochroleuca [vajszínű ördögszem]) és/vagy színes fellevelek kapcsolódnak hozzájuk (pl. Araceae [kontyvirágfélék], Euphorbiaceae [kutyatejfélék] családjai). A csalogató színezet nem feltétlenül az emberi szem számára érzékelhető hullámhossz-spektrumba esik, a fehérnek tűnő virágokon gyakran az UV-tartományba eső mintázat található. A trópusokon élő madárporozta virágok jellemzője, hogy erőteljes (többnyire vöröses, narancsos) színezetük ellenére nem illatosak.
A zoogámiát szolgáló szervezettani jellemzők
A növényvilágban viszonylag ritka jelenség a víz segítségével történő megporzás (hidrogámia). E fajok virágainak virágtakarója is jelentéktelen, redukált. A víz alatt nyíló fajok (pl. Ceratophyllum [tócsagaz]) pollenszemei a vízzel azonos fajsúlyúak, gyakran fonal alakúak (és nincs exinéjük). A vízfelszínén nyíló virágokkal rendelkező nemzetségekben (pl. Vallisneria [csavarhínár], Anacharis [átokhínár]) gyakori, hogy a növényről leváló porzós virágok teljes egészükben sodródnak el a termős virágokhoz.
A megporzástól a kettős megtermékenyítésig
A bibe felületére került pollenszem vegetatív sejtje – fogadóképes bibe esetében – a bibe cukros váladékának hatására ozmotikus vízfelvétellel megnöveli belső turgorát, aminek az lesz az eredménye, hogy a sejtfal intine rétege kitüremkedik a pollenfal valamelyik pórusán vagy hasítékán, és a pollen kicsírázik (9/a ábra). A virágporszemből kihajtó pollentömlő (csíratömlő) valójában nem más, mint a vegetatív sejt állábhoz hasonló citoplazma-nyúlványa, ami a bibe, majd a bibeszál szöveteibe hatolva halad a magkezdemények felé. A vegetatív sejt magja és a generatív sejt is belép a pollentömlőbe, és annak plazmájában úszik a magház felé. A bibeszál vezető szövetében haladó csíratömlő pollenszem felé eső szakaszai kallózból álló sejtfalakkal („kallózdugó”) lezáródnak, majd degenerálódnak. Binukleátus virágporszemek esetében a generatív sejt a magkezdemények felé haladása közben mitotikusan osztódva létrehozza a két hímivarsejtet. (A trinukleátus pollenben magukat a hímivarsejteket találjuk, tehát közvetlenül ezek lépnek be a csíratömlőbe.)
A megtermékenyítés segítése: a szinergidák feladata
A magház üregébe jutó csíratömlő a magház falának belső oldalán halad a placenta felé, ahonnan a köldökzsinóron át eljut a magkezdeményig. Az esetek többségében a mikropile felől belenő a magkezdemény állományába, átjut a nucelluszon az embriózsákig. Ekkorra a központi sejt diploid magja felúszik a petesejt-apparátus közelébe. A pollentömlő belenő az egyik segítősejtbe (szinergidába), ami addigra apoptotikus folyamatban degradálódni kezd (9/b ábra). A pusztuló szinergida belső állományának hatására a pollentömlőt ozmotikus sokk éri, ráadásul a szinergida autolízise számos sejtfalbontó hatású enzim megjelenését eredményezi, ami elvékonyítja a tömlő falát. Ez lehetővé teszi a hímivarsejtek kiszabadulását.
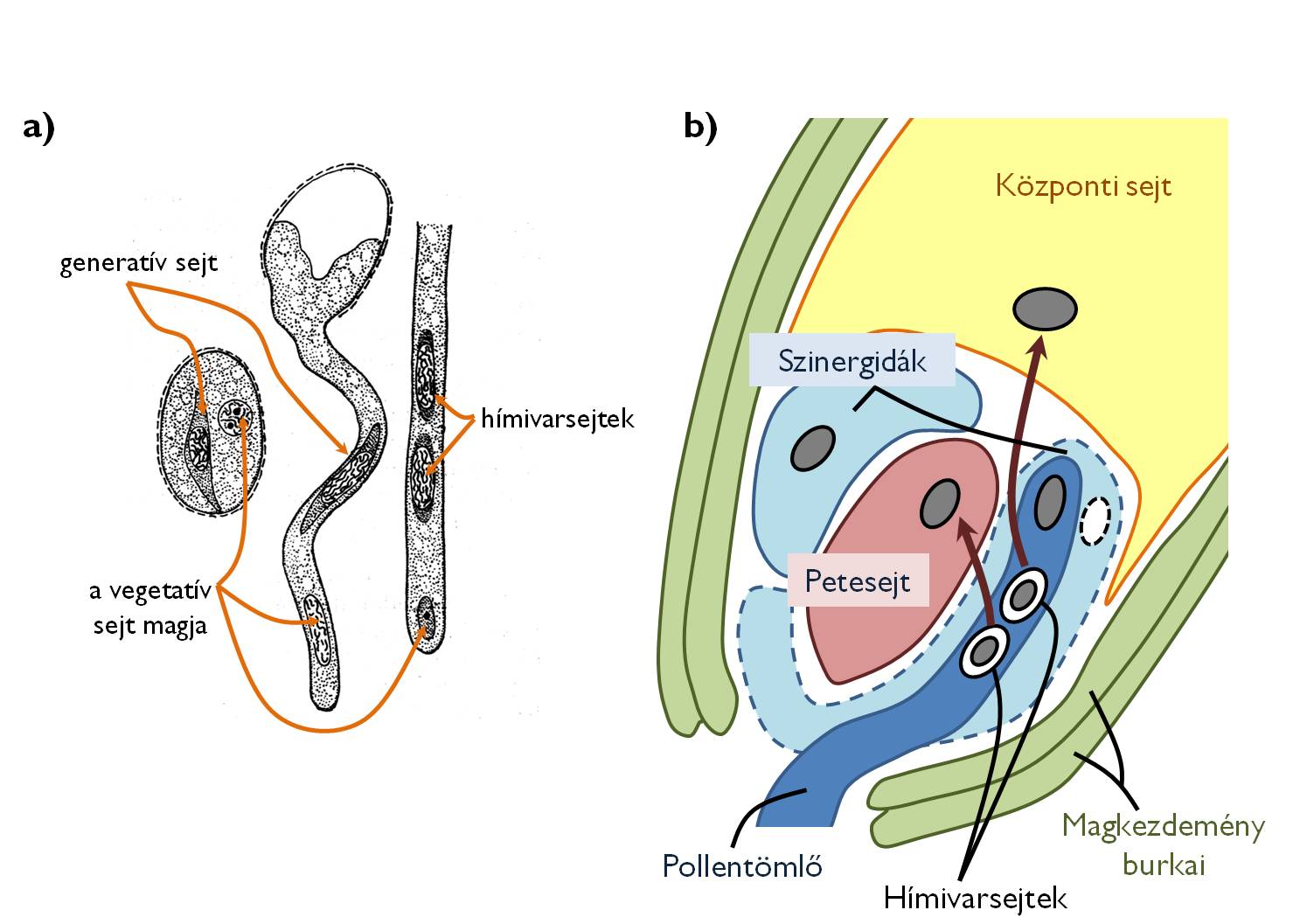
9. ábra. A pollentömlő kialakulása (a) és a kettős megtermékenyítés (b).
(a: Kárpáti nyomán)
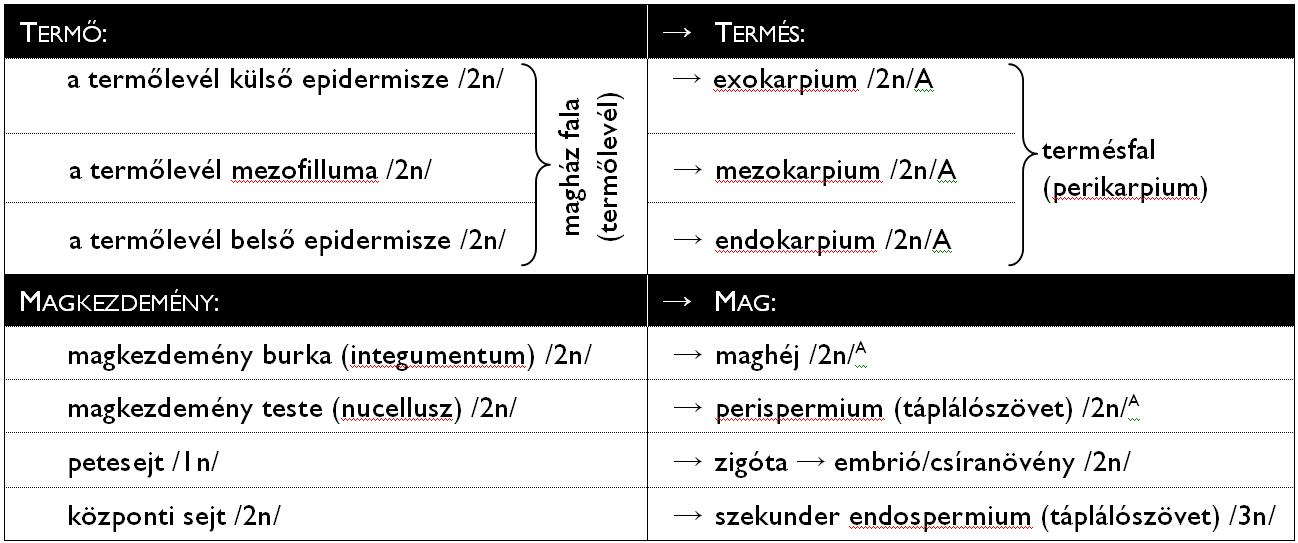
A kettős megtermékenyítés (amfimixis) eseménye során az egyik hímivarsejt a haploid petesejttel, míg a másik a diploid központi sejttel olvad össze. A folyamat eredményeként jön létre a petesejtből a diploid (2n) zigóta, ami hamarosan embrióvá (majd csíranövénnyé) fejlődik, a központi sejtből pedig kialakul a triploid (3n) szekunder endospermiumnak nevezett táplálószövet. A megtermékenyítés után a magkezdemény burkaiból (integumentumok) kialakul a maghéj, a magkezdemény nucelluszából pedig a perispermiumnak nevezett, anyai génállományú táplálószövet. A magkezdemény ezzel maggá alakul át (4. táblázat). (Megjegyzendő tehát, hogy a magot körülvevő maghéj és a perispermium az anyai sporofitonnal megegyező információtartalmú részletek.) A megporzástól a megtermékenyülésig, zárvatermők esetében (a nyitvatermőkkel szemben) mindössze néhány óra telik el.
4. táblázat. A kettős megtermékenyítéssel végbemenő változások.
(A zárójelben szereplő számok a ploidiaszintet jelölik, az A-val jelölt részek génállománya megegyezik az anyai sporofitonéval.)
A termés
A termés csak a zárvatermőkre jellemző szaporítószerv, amelynek elsődleges feladata a benne levő magok fejlődés alatti védelme valamint későbbi terjesztése.
A megtermékenyítést követően a virág termőjének szövetei átalakulnak: a magház falának szerkezete megváltozik, termésfallá (perikarpium) alakul (10. ábra). A termőlevelek szöveti felépítésének megfelelően a termésfalnak három jellemző rétege van: a termőlevél külső epidermiszéből kialakuló exokarpium, a mezofillum átalakulásával létrejövő mezokarpium és a magház ürege felé eső epidermiszből fejlődő endokarpium. Számos növényfaj esetében a magház fala körül – a fokozott magvédelem céljából – egyéb virágrészek (vacok, takarólevelek alapi részei stb.) is összezáródhatnak. Ekkor áltermésről beszélünk, megkülönböztetve ezeket az eseteket a csak a termőből kialakuló valódi termés ektől.
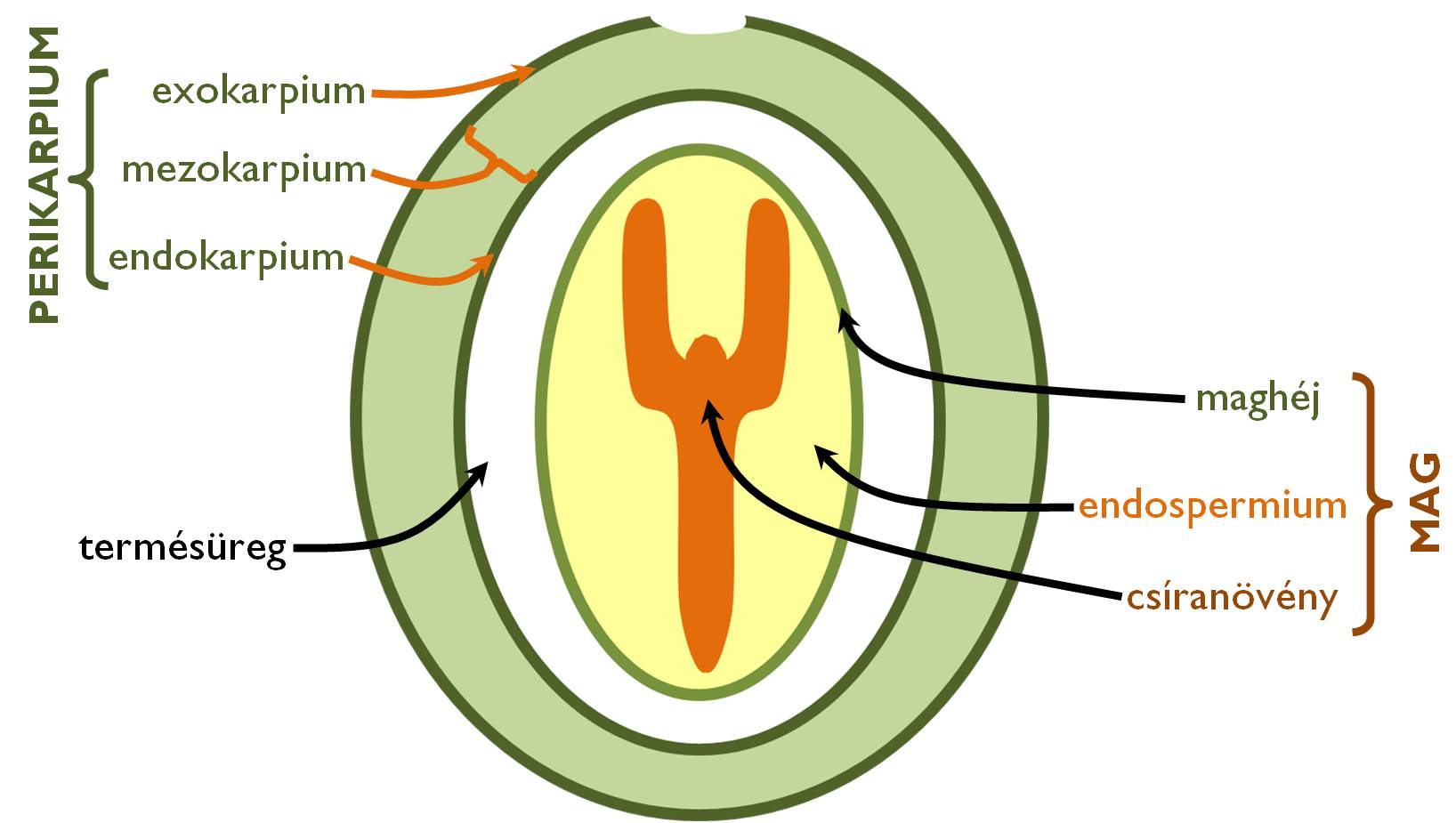
10. ábra. A termés és a mag felépítése.
(A zölddel szedett részek az anyai génállományú, diploid szövetek!)
Termésfejlődés megtermékenyítés nélkül: a partenokarpia
A virágok termőinek térbeli viszonya befolyásolja a termések csoportosulását. Ha a terméskocsányon egyetlen termés helyezkedik el (ami mono- vagy cönokarp magházból jött létre), magányos termésről beszélünk (pl. Syringa vulgaris [közönséges orgona], Aesculus hippocastanum [fehér bokrétafa]). Ha a virágnak több termője van (azaz termőtája apokarp), akkor egyetlen kocsányon több termés helyezkedik majd el: csoportos termés (pl. Clematis vitalba [erdei iszalag], Rubus idaeus [málna]). Az is előfordulhat, hogy egy kocsányon több olyan termés található, amelyek külön virágokból (egy virágzat tagjaiból) alakultak ki. Később ezek többé-kevésbé összenőnek egymással, így együtt terjednek. Ez a terméságazat, melynek kialakításában a virágzati tengely és a virágok egyéb részei is részt vehetnek (pl. Ananas comosus [ananász], Ficus carica [füge], Morus alba [fehér eperfa]).
Attól függően, hogy a magterjesztés állapotában a termésfal alacsony vagy magas nedvességtartalmú-e, megkülönböztetünk száraz és húsos terméseket. Ha a magterjesztés állapotában a termésfal valamennyi rétege felnyílik, a termést felnyílónak mondjuk, ha a termésfal nem (vagy nem minden rétege) nyílik fel, a termés zárt termés. A zárt termés speciális esete a hasadó termés, amikor a magterjesztés állapotában a termésfal nem nyílik fel, de a termés egymagvú résztermésekre esik szét (pl. a Lamiaceae [ajakosok] és a Boraginaceae [érdeslevelűek] családokra jellemző négy makkocska termés vagy az Apiaceae családban [ernyősök] általános ikerkaszattermés).
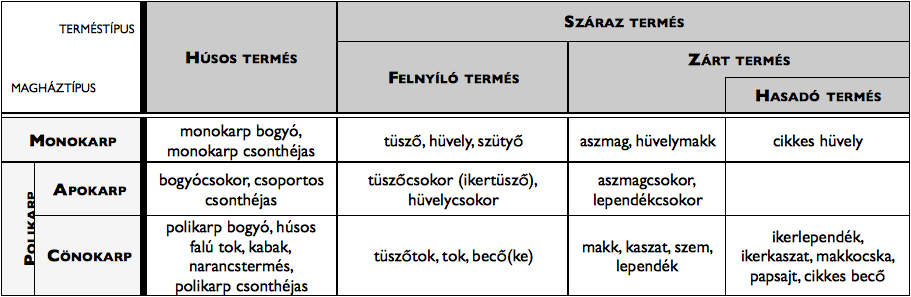
Mivel az egyes terméstípusok áttekintése elsősorban alaktani kérdés, részletes leírásuk helyett (melyet a morfológiai szakirodalom tartalmaz) csak a különböző típusok áttekintő rendszerét közöljük a 5. táblázatban.
5. táblázat. A terméstípusok áttekintése.
A termés szövettana
Változatos kialakulásuk miatt a termések (és áltermések) szövettana típusonként (sokszor még azon belül, rendszertani csoportonként is) változik. Ennek ellenére néhány általános jellemző elmondható a perikarpium egyes rétegeinek szerkezetéről.
Az exokarpium szinte minden esetben egyrétegű epidermisz, amelyen sztómák, gyakran növényi szőrök (fedőszőrök, serteszőrök, akaszkodó szőrök), vastagabb kutikula és/vagy viaszréteg található. E réteg érdekessége, hogy a termésfejlődés során viszonylag sokáig megőrzi merisztematikus aktivitását, sejtjei antiklinális falakkal folyamatosan osztódnak.
A mezokarpium a termésfal legváltozatosabb anatómiájú rétege. Benne minden esetben redukált kollaterális zárt szállítónyalábok (a termőlevél erei) futnak, amelyek a termésfal szöveteinek táplálásában vesznek részt. Húsos termések esetében fő tömegét raktározó parenchima alkotja (terméshús, pulpa), benne felhalmozott tápanyagokkal, szerves savakkal, cukrokkal, színanyagokkal, speciális (másodlagos) anyagcseretermékekkel. Száraz termések esetében állománya vékonyabb, nem ritkán szklerenchimatikus rétegek teszik ellenállóvá (pl. a makktermés esetében). (Szilárdító alapszöveti rétegek a húsos falú termések mezokarpiumában is előfordulhatnak.) Felnyíló termések esetében többnyire ebben a rétegben találjuk a termés felnyílásáért felelős szöveti elemeket is, amelyek a vízvesztés okozta zsugorodása nyomán fellépő húzóerő révén felrepesztik a perikarpiumot.
Az endokarpium a termések többségében egyrétegű, módosult epidermiszként jelenik meg. Mivel azonban az általános bőrszöveti funkciók ellátására ebben az esetben nincs szükség, szerkezete sok esetben módosult az evolúció során. Sejtjei parenchimatikussá alakulhatnak és részt vehetnek a terméshús kialakításában (pl. a bogyótermés esetében). A csonthéjas termés endokarpiumát kősejt-rétegek alkotják; ez a csontkeménységű csonttár (drupa) védi a benne levő magot. Az endokarpium a termések egy része esetében (pl. szemtermés, ikerkaszat) összenő a maghéjjal.
A termésfal többnyire a magház üregéből kialakult, magvakat tartalmazó üreget (termésüreg) zárja körül. Ezt kitöltheti levegő (pl. Capsicum annuum [paprika]) vagy folyadék (pl. Solanum lycopersicon [paradicsom]), de kitöltheti az endokarpiummal kapcsolatban álló laza szövet („termésbél”) (pl. Citrullus lanatus [görögdinnye]) vagy akár az endokarpium epidermális eredetű szőrei (pl. Citrus sinensis [narancs]) is.
A mag
A mag a nyitvatermők ősi csoportjaiban (a magvaspáfrányok körében) megjelent evolúciós újítás. A benne található embrió védelmére, táplálására és – a nyitvatermők valamint a felnyíló termésű zárvatermők esetében – terjesztésére szolgáló szaporító képlet. Az anyanövény testén (az anyai sporofitonon) alakul ki a magkezdeményből, a (kettős) megtermékenyítés eredményeként. Leválását követően hormonális (abszcizinsav) hatásra bekövetkező, hosszabb vagy rövidebb nyugalmi időszak (magnyugalom) jellemzi, ami a csírázás megindulásával szakad meg.
A mag felépítése
A magvakat minden esetben a magkezdemény burkából (az integumentumból) létrejövő maghéj borítja. Ez a fajok többségénél kétrétegű: külső rétege a teszta, belső rétege a tegmen. A köldökzsinórról történő leválását követően jellegzetes folt, a köldök (hilum) vagy köldökfolt válik láthatóvá rajta, ami egyes esetekben (pl. Fabaceae család tagjai [hüvelyesek], Aesculus hippocastanum [fehér bokrétafa], Cardiospermum halicacabum [szívmag]) jellegzetes alakú, méretű. A hilum körül egyéb képletek is megfigyelhetők. Ha a köldökzsinór hosszabban ránő a mag felületére (pl. anatrop magkezdemény esetében), akkor egy varrat (rafe) látható a maghéjon. Hangyák terjesztette magokon egy dudorszerű kinövés, a hangyakenyér (karunkula, elaioszóma) található a köldök körül, amiben a terjesztő rovarok számára értékes tápanyagok (olajok, fehérjék) halmozódnak fel.
A maghéj hordozhat különböző függelékeket is, amelyek többnyire a terjedést segítik. Ilyenek a magszőrök (pl. Salix [fűz], Gossypium [gyapot]) vagy a magszárnyak (pl. Pinus [fenyő] fajok). A maghéjon kívül egyes magokon feltűnő színű, húsos magköpeny (arillusz) is kialakulhat, ami nem az integumentum, hanem a köldökzsinór származéka. Magköpeny segíti pl. a Taxus baccata [tiszafa], az Euonymus [kecskerágó] vagy a Paeonia [pünkösdirózsa] fajok magvainak terjedését, de előfordul a Nymphaeaceae [tündérrózsafélék] család képviselőinek magjain is. A szerecsendió (Myristica fragrans) piros színű, hálózatos arillusza – mint „szerecsendió-virág” – fűszerként kerül az üzletekbe.
A maghéj legkülső rétegét egyszerű vagy módosult epidermisz borítja. Az epidermális képletek (kutikula, szőrök) mellett nyálkaanyagot felhalmozó sejtek is előfordulhatnak benne (pl. Linum [len], Sinapis [mustár]). A belsőbb, parenchimatikus rétegek között szilárdító elemek (szklereidák, rostok) fordulhatnak elő, különösen a felnyíló termésekből kiszóródó magvak esetében. Néha erősen redukált szállítónyalábok is felismerhetők benne. Ritkán húsos, plazmadús parenchima is alkothat egyes rétegeket, mint pl. a Ginkgo biloba [ginkgófa] magjának külső szarkotesztája esetében (a faj maghéjának belső rétege kősejtekből álló szkleroteszta).
A mag belső állományát az embrió (csíranövény) és a táplálószövet alkotja. Az embrió táplálására szolgáló raktározó parenchima különböző eredetű lehet. (+info) A nyitvatermők magvaiban a női gametofiton haploid sejtjeiből kialakuló primer endospermium (1n) a fő táplálószövet. A zárvatermők magjaiban ezt a szerepet elláthatják az embriózsák központi sejtjéből – a kettős megtermékenyítés eredményeként – kialakuló szekunder endospermium sejtjei (3n), vagy akár a magkezdemény anyai szöveteinek (nucellusz) leszármazottai (perispermium) (2n) is. Vannak mindazonáltal olyan zárvatermő fajok is, amelyek magjai egyik felsorolt táplálószövet-típust sem tartalmazzák. Ebben az esetben a sziklevelek (vagyis az embrió diploid szövetei) akkumulálnak nagyobb mennyiségű tápanyagot. A zárvatermőkre jellemző táplálószövetek aránya fajonként változó lehet, néha többféle is előfordulhat egy-egy magban. Az egyes tápanyag-raktározási módokra mégis adhatunk egyértelmű példákat is:
-
- primer endospermiumos magvak: nyitvatermő fajok;
-
szekunder endospermiumos magvak: Poaceae [pázsitfűfélék], Liliaceae [liliomfélék], Solanaceae [burgonyafélék], Apiaceae [ernyősök], Polyagonaceae [keserűfűfélék] családok;
-
perispermiumos magvak: Caryophyllaceae [szegfűfélék] számos nemzetsége;
-
szekunder endospermiumot és perispermiumot is tartalmazó kettős tápszövetű magvak: Piperaceae [borsfélék], Cannabidaceae [kenderfélék], Amaranthaceae [disznóparéjfélék], Zingiberaceae [gyömbérfélék] családok;
-
raktározó szikleveles magvak: Fabaceae [hüvelyesek], Fagaceae [bükkfafélék], Rosaceae [rózsafélék], Asteraceae [fészekvirágzatúak] családok, Cucirbitaceae [tökfélék].
Embriófejlődés megtermékenyítés nélkül: az apomixis
A termések és magvak terjedése
A termés és a mag kialakulásának legfontosabb célja a szaporodás és a terjedés. Annak érdekében, hogy ezt a feladatukat teljesíthessék, fontos, hogy a bennük levő utód (a csíranövény) kisebb-nagyobb távolságra eljusson az anyanövénytől. A magvak és termések terjedési lehetőségei alapvetően meghatározhatják egy növényfaj terjedési képességeit, ezáltal földrajzi (és ökológiai) elterjedési területét. A növények evolúciója során különböző módok alakultak ki a propagulum-terjedés elősegítésére, amelyek felnyíló termések esetében zömmel a mag, zárt termések esetében a termés(fal) anatómiai vagy morfológiai megváltozásával jártak.
Az anemochoria a széllel, légmozgással történő terjedést jelenti. A virágos növények között viszonylag alacsony az olyan fajok száma, amelyek felnyíló terméseiből olyan apró, könnyű magvak kerülnek a külvilágba, amelyek kis fajsúlyuknál fogva nem igényelnek speciális képleteket terjedésükhöz (pl. Orchidaceae család [kosborfélék], Petunia [petúnia], Begonia [begónia], Orobanche [vajvirág] fajok). Gyakoribb eset, hogy a szél vagy az egész növényt (pl. Eryngium [iringó, ördögszekér], Salsola [ballagófű]) vagy annak egy részét (pl. Papaver [mák]) leválasztja, görgeti és e mozgás során szóródnak ki a magvak a talajra. Nagyobb termések illetve magvak esetében már speciális repítőkészülékekre lehet szükség. Ezek lehetnek különféle „szárnyak”, amelyek akár a mag (pl. fenyőfélék, Campsis [trombitafolyondár], Catalpa [szivarfa]), akár a termésfal (lependéktermés – Fraxinus [kőris], Ulmus [szil]; ikerlependék termés - Acer [juhar]) függelékei lehetnek, de fellevelek is elláthatják funkciójukat (pl. Tilia [hárs], Carpinus [gyertyán]). Az anemochoriát segíthetik szőrképletek is. Ezek szintén előfordulhatnak a magon (pl. Gossypium [gyapot], Asclepias [selyemkóró], Salix [fűz], Populus [nyár]), vagy csatlakozhatnak a terméshez. Bibeszál-eredetű szőrképletek csatlakoznak a Clematis [iszalag] vagy a Pulsatilla [kökörcsin] fajok aszmagterméseihez, tollas szálkájú toklász nő az árvalányhajak (Stipa) szemtermésére, a szőrszerűen redukálódott csészelevelekből kialakulóbóbita (pappusz) repíti egyes fészekvirágzatúak kaszattermését (pl. Taraxacum [pitypang], Tragopogon [bakszakáll]). A Cotinus coggygria [cserszömörce] csonthéjas terméseinek terjedését a bugavirágzat steril virágainak szőrössé váló kocsányai segítik.
A vízzel terjedő (hidrochor) termések jellemzője, hogy falukon a víz nem hatol át, ugyanakkor a termésüreget levegő (vagy a víznél kisebb fajsúlyú folyadék) tölti ki (pl. Nuphar [vízitök], Nymphaea [tündérrózsa], Trapa natans [sulyom]). A vízfelszínen lebegő terméseket ekkor az áramlás viszi tova, akár több 100 kilométeres távolságra is (pl. Cocos nucifera [kókusz], Lodoicea seychellarum [maldív dió]).
A propagulumok terjesztésében állatok is részt vehetnek (zoochoria), méghozzá többféle módon is. Az endozoochoria esetében a húsos (vagy ritkán a száraz) termést az állat elfogyasztja, de tápcsatornáján a magvak sérülés nélkül keresztülhaladnak és ürítés után csíráznak. Ezen fajok termései, gyümölcsei feltűnő színűek, magas tápanyagtartalmúak, illatosak és gyakran nagy méretűek (pl. Vitis [szőlő], Rubus [szeder], Malus [alma]). Egyes endozoochor növények magvai nem is képesek kicsírázni mindaddig, amíg a terjesztő állat bélcsatornájának megfelelő enzimei fel nem lazítják maghéjukat. Ennél gyakoribb eset, amikor a termés vagy a mag akaszkodó szőrökkel, horgokkal (pl. Arctium [bojtorján],Xanthium [szerbtövis], Hibiscus [mályvarózsa], Galium aparine [ragadós galaj]) vagy enyves váladéka segítségével (pl. Viscum [fagyöngy], Linum [len]) ráakaszkodik a terjesztő állat bundájára, tollazatára (epizoochoria). A termések és magvak egy részét az állatok elhordják, összegyűjtik, s miközben elhullajtják azokat, elvégzik terjesztésüket is (szinzoochoria). Ezt a terjesztési munkát igen gyakran hangyák végzik (mirmekochoria), s ebben az esetben a mag felületén találunk egy fehérjékben és/vagy olajokban gazdag köldökpúpot (karunkula, elaioszóma), ami tápanyagot szolgáltat az állatok számára (pl. Viola [ibolya], Chelidonium [fecskefű], Corydalis [keltike], Ricinus [ricinus]).
Külön csoportot alkotnak az ún. dinamochor („magrúgó”) termések, amelyek esetében a termés maga hordoz a magvak terjesztésére szolgáló szervi módosulatot. Rendszerint a felnyíló termésfal mozgása szolgáltatja a magok kiszórásához szükséges energiát (pl. a hüvelytermés vagy az Impatiens [nenyúljhozzám], Viola [ibolya] vagy Oxalis [madársóska] nemzetségek toktermései esetében). A magrúgó (Ecballium) termése a magszóráskor hirtelen leválik kocsányáról, mire a belsejét kitöltő nagy nyomású folyadék a magvakkal együtt messzire spriccel belőle.
Az embriogenezis és a csírázás folyamatai
Az embriófejlődés folyamata (embriogenezis)
A szárazföldi növények (Embryophyta) közös bélyege, hogy zigótájukból nem közvetlenül a növény, hanem először egy zömmel merisztematikus sejtekből álló embrió fejlődik ki. Az embriogenezis folyamata rendszertani csoportonként eltérő, ezért jelen fejezetben elsőként a zárvatermők törzse (Magnoliophyta) valódi kétszikűek osztályára (Rosoposida) jellemző alapszabást tárgyaljuk, majd számba vesszük az ehhez képest megfigyelhető eltéréseket.
A zigóta sejtjének osztódásai képesek létrehozni a teljes növényi szervezetet. Leszármazottaiból bármely növényi szerv minden szövete kialakulhat. Az ilyen sejteket nevezzük totipotens sejteknek. A totipotens sejtek utódsejtjei egyre differenciáltabbakká válnak, elköteleződnek egyes szervek iránt, azaz mitózisaikkal már csak egy adott növényi szerv szövetei jöhetnek létre. Ezek az ún. pluripotens sejtek. A szervkezdemények sejtjeinek mitózisaival végül olyan – még mindig osztódóképes – sejtek jönnek létre, amelyek már szöveti szinten elkötelezettek, azaz belőlük már csak az adott szerv egy meghatározott szövetrendszerének sejtjei alakulhatnak ki. Ezen merisztematikus sejteket multipotensnek mondjuk. A növényi szervezet érdekessége, hogy hormonális hatásra a sejtek elkötelezettségi szintje megváltozhat: a multipotens sejtek pluripotenssé, vagy akár totipotenssé válhatnak (sérülések vagy vegetatív szerveken létrejövő embriók kialakulását megelőzően).
Az embriogenezis folyamata az embriózsákban létrejött zigóta inekvális osztódásával indul (11. ábra). Ennek eredményeként egy kisebb méretű, sűrű citoplazmájú csúcsi (apikális) sejt és egy nagyobb méretű, erősen vakuolizált alapi (bazális) sejt jön létre. Ezzel a lépéssel a proembrió polarizálttá válik: az apikális sejtből fejlődik ki majd az embrió nagy része, míg az alapi sejt nagyrészt a csírafüggesztő (szuszpenzor) kialakításában vesz majd részt. A kétsejtes proembrió alapi sejtje ezt követően ismét inekválisan osztódik a hossztengelyre merőleges, transzverzális fallal. Alsó, nagyméretű sejtje a szuszpenzorsejt, amelynek későbbi transzverzális osztódásai alakítják ki a csírafüggesztőt, ami az embriót rögzíti a magkezdemény mikropile felé eső oldalán illetve tápanyagokat (és fejlődést szabályozó anyagokat) közvetít az anyanövény és az embrió között. Az alapi sejt felső, apikális sejt felé eső utóda a hipofízissejt, ami hosszanti falakkal (longitudinálisan) osztódik négyfelé. A későbbiekben ezek a sejtek a proembrió gyökérpólusának (gyököcskéjének) a kialakításában vesznek majd részt.
A kétsejtes proembrió apikális sejtje kétszer egymás után az embrió hossztengelyével párhuzamosan (longitudinálisan) osztódik, létrehozva a kvadráns (négysejtes) állapotot. Ezután a négy sejt transzverzális fallal osztódik tovább oktánssá. Az oktáns sejtjei ekkor periklinális osztódásokkal gyarapítják tovább a proembrió tömegét, ezzel annak merisztematikus sejtjei egy kerületi (felszíni) és egy központi részre különíthetők. Ebben az időszakban a proembrió gömb alakot vesz fel (gömb stádium). A felszíni sejtek antiklinális osztódásokkal fokozatosan az epidermiszt létrehozó protodermává alakulnak, míg a központi zónában peri- és antiklinális osztódások egyaránt jellemzőek. A központi zóna felszíni részén erősebben vakuolizált sejteket találunk, amelyekből a későbbi alapmerisztéma jön létre, míg a közbülső, fokozatosan tengelyirányba rendeződő, kevésbé vakuolizált sejtek a prokambium előfutárai.
A következő időszakban a gömbstádiumú proembrió sejttömegének belsőbb régióiban két helyen élénk periklinális osztódásokkal megindul a sziklevelek fejlődése. Később ezek a proembrió felszínén kidudorodásokat képeznek: létrejön a szív stádium. A sziklevelek további fejlődésével és az embrió tengelyének (lényegében hipokotiljának) megnyúlásával ebből alakul ki a torpedó stádium. Ezzel párhuzamosan a korábban az egész proembrióra jellemző mitotikus osztódások mindinkább csak a két pólusra korlátozódnak. A sziklevelek közti területre, az embrionális hajtáscsúcsra (rügyecske, plumula) valamint a proembrió szuszpenzorral érintkező, a hipofízissejt osztódásaival létrejött régiójára, az embrionális gyökércsúcsra (gyököcske, radikula). (+info) A csíranövény sziklevelek alatti (gyökérnyakig tartó) szárrésze a hipokotil, míg a sziklevelektől az első, embrionális állapotú levelekig tartó szárrész az epikotil.
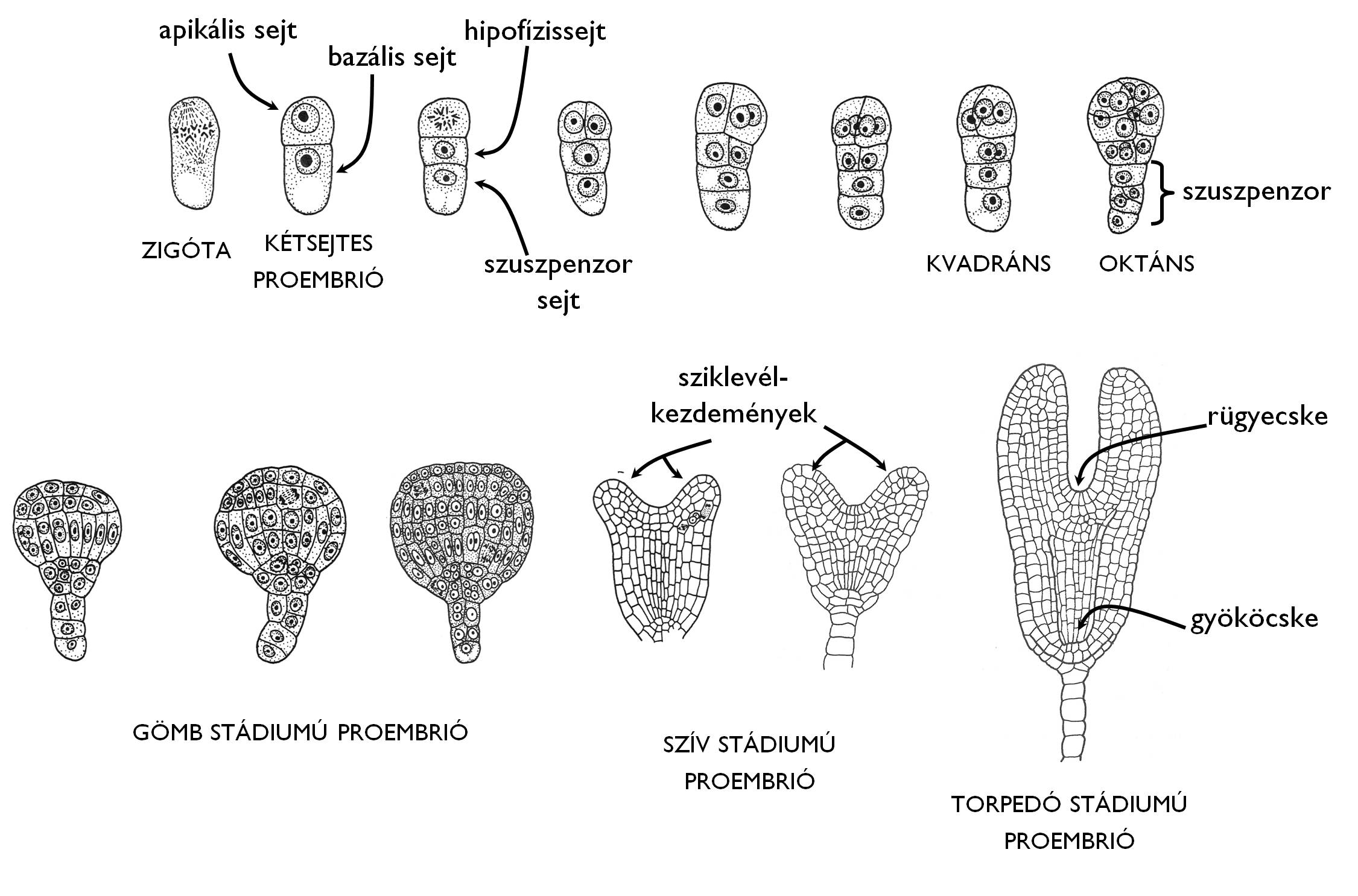
11. ábra. A valódi kétszikűek (Rosopsida) embriogenezisének alapvonásai.
(Gyurján nyomán)
Az egyszikűek embrigenezisének sajátosságai
A nyitvatermők embrigenezisének sajátosságai
A csírázás
A zigótából kialakuló embrió a harasztok esetében nyugalmi állapot nélkül fejlődik tovább, és az előtelepen kialakul belőle a harasztnövény. A magvas növények esetében ezzel szemben az utód sporofitonjának (a csírának) a fejlődése a polarizálódás, az embrionális hajtás- és gyökércsúcs valamint a sziklevelek kialakulása után leáll: beáll a magnyugalom állapota.
A magnyugalom megtörésére a csírázás megindulásával kerül sor. A növények csíraképessége számos külső és belső tényezőtől függ. A sikeres fejlődéshez érett és ép magvakra van szükség, ami nem feltétlenül áll összefüggésben a termés fejlettségével. (Az érettnek tűnő őszibarack termésében például csak igen későn válik csíraképessé a mag, míg a borsó vagy bab még zöld színű termései már érett magvakat tartalmaznak.) Nyugalmi állapotban a magvak többsége a szélsőséges körülményeket (hideg, meleg, szárazság, oxigénhiány) is elviseli, ami előnyösen szolgálja a magvak hosszú időn át tartó, nagy távolságokat áthidaló terjedési folyamatait.
A csírázás megindulásának külső feltételei a megfelelő ökológiai körülményeket jelentik, elsősorban a felvehető vizet és az optimális hőmérsékletet. A melegebb égövi növények többsége magasabb hőmérsékleten csírázik, mint a sarkokhoz közelebbi területekről származó fajok magvai. Számos mérsékelt övi növény magjának csírázáshoz tartósabb hideg- vagy fagyhatásra van szükség. A megvilágító fény mennyisége is hatással lehet a csírázásra. A pázsitfűfélék magjai például csak fényen indulnak fejlődésnek.
Számos csíranövény megfelelő fejlődéséhez más élőlények hatására van szükség. Az orchideafélék vagy a nyitvatermők többségének csíranövénye csak abban az esetben éli túl a csírázást követő első néhány napot, ha megfelelő gombapartnerrel találkozva képes kialakítani a számára létfontosságú mikorrhiza-kapcsolatokat. Egyes vastagabb maghéjú magok csak növényevők tápcsatornáján áthaladva válnak csíraképessé.
A növényi magvak csíraképességének időtartama változó. Rövid ideig (egy hét) csíraképesek például a fűz- és a nyárfajok magvai. Néhány évig tartó nyugalmi állapotot is elviselnek a legtöbb zöldségnövényünk magjai, míg a gyommagvak akár 40 év múltán is fejlődésnek indulhatnak. A csírázási képesség hosszabb ideig történő megőrzése szintén megfelelő ökológiai stratégia lehet a hosszabb távra történő terjedés vagy a tartósan kedvezőtlen időszak átvészelésére. Ez a magyarázata annak is, hogy egyes területeken a talajban található magkészlet összessége (magbank) nem feltétlenül felel meg a területen hajtásokkal is előforduló növényi fajkészletnek. Ennek ellenére a magbankban benne rejlik a fajösszetétel megváltozásának lehetősége a környezeti feltételek változása esetén.
A csírázás folyamata a magvak passzív vízfelvételével, a duzzadással indul meg. Ennek során a vizet áteresztő maghéjon keresztül ozmózissal nagy mennyiségű vizet vesznek fel az addig mindössze 10-15% nedvességet tartalmazó magvak. A magba jutó víz hatására a dehidratált állapotban levő enzimek aktiválódnak és a magban beindulnak a lebontó anyagcsere-folyamatok, köztük a tartaléktápanyagok lebontása. A táplálószövetekben felhalmozott szervesanyag lebontása elegendő anyagot és energiát termel ahhoz, hogy megindulhasson a csíranövény fejlődése. Az aktiválódó csúcsmerisztémák működésbe lépésével először a fiatal gyökér, majd a hajtás is kilép a magból, megindulhat a csíranövény önálló tápanyagfelvétele, majd a hajtás zöldülésével a fotoszintézis is.
A csíranövény fejlődése során jellemző, hogy a sziklevelek a talajfelszín alatt (a maghéjon belül) maradnak-e, vagy a maghéjból kiszabadulva átlépik a talajfelszínt, és ott megzöldülve fotoszintézisbe kezdenek-e. Az előbbi esetben föld alatti (hipogéikus), az utóbbi esetben föld feletti (epigéikus) csírázásról beszélünk. A kétféle csírázási mód nem feltétlenül köthető rendszertani csoporthoz. Hipogéikus csírázású fajok pl. a Poaceae család [pázsitfűfélék] tagjai, a Phaseolus coccineus [tűzbab], az Aesculus hippocastanum [fehér bokrétafa], a Prunus domestica [szilva] vagy a Quercus robur [kocsányos tölgy], míg epigéikus csírázásúak pl. a Brassicaceae [keresztesvirágúak], Asteraceae [fészekvirágzatúak], Cucurbitaceae [tökfélék], Pinaceae [fenyőfélék] családok vagy az Allium [hagyma] nemzetség tagjai.
Ellenőrző feladatok
Ide jó lenne a feladatokat feltölteni
Irodalomjegyzék
Erős-Honti Zs (2012): A kertészeti növények alaktana. Egyetemi jegyzet. Budapesti Corvinus Egyetem Kertészettudományi Kar, Bp.
Gyurján I (1996): Növényszervezettan. Egyetemi jegyzet. ELTE Eötvös K., Bp.
Haraszty Á (1998): Növényszervezettan és növényélettan. 6. kiad., Nemzeti Tankvk., Bp.
Jacob F, Jäger EI, Ohmann E (1985): Botanikai kompendium. Natura, Bp.
Kárpáti Z, Görgényi L, Terpó A (1968): Kertészeti növénytan I. Növényszervezettan. Mezőgazdasági Kiadó, Bp.
Mauseth JD (2008): Plant Anatomy. The Blackburn Press, New Jersey.
Mishra SR (2009): Understanding Plant Reproduction. Discovery Publishing House Pvt. Ltd., New Delhi.
Sárkány S, Haraszty Á (1995): Növényszervezettan biológia-kémia, biológia ill. pszichológia-biológia szakos hallgatók részére. Egységes jegyzet. Nemzeti Tankvk., Bp.
Sárkány S, Szalai I (1966): Növénytani praktikum I. Növényszervezettani gyakorlatok. 3. kiad., Tankvk., Bp.
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028