A szervek szövettana (Organográfia)
Szerző: Erős-Honti Zsolt
A gyökér
A gyökér evolúciós eredete és kialakulása, tagolódása és architektúrája
A növényi gyökérzet elsődleges szerepe a tápanyagok (víz és ásványi sók) felvétele a talajból, a növényi szervezet rögzítése valamint évelő növények esetében a kész szerves anyagok raktározása a vegetációs időszakot követő nyugalmi állapot során, ezzel fedezve a következő tenyészidőszak elején kihajtó, fotoszintézisre egy ideig képtelen növény tápanyagigényét.
Annak ellenére, hogy a gyökér a növény egyedfejlődése során legkorábban megjelenő szerv, evolúciós szempontból éppen a legfiatalabb. Kialakulásának hátterében az állt, hogy a szárazföldön a tápanyagok felvétele csak a talajból történhetett meg s nem a teljes testfelszínen keresztül, így szükség volt egy olyan felszívó szervre, ami képes a korlátozottan hozzáférhető anyagok hatékony felszívására. Az elsőként kialakult hajtásos növénycsoport tagjainak, a harasztoknak csak hajtás eredetű, járulékos gyökereik vannak, mivel embriójuknak csak egyetlen osztódó pólusa (rügyecskéje) van (unipoláris embrió), amelyből a hajtás alakul ki. A gyököcskével (radikula) rendelkező bipoláris embrió és a valódi gyökérzet elsőként az ősi nyitvatermők körében jelent meg, tehát csak a hajtás kialakulását követően.
Valódi gyökérzetnek a kétpólusú (bipoláris) embrió gyököcskéjéből (radikula) eredeztethető gyökérrendszert nevezzük, megkülönböztetve azt a hajtás eredetű gyökérzettől. Valódi gyökérzet esetében a radikula eredetű gyökérág fejlődése során domináns szerepű főgyökérré válik, amelynek elágazódásával alakulnak ki az oldalgyökerek (ez a főgyökérzet, vagy allorhiza). A hajtás eredetű gyökérzet egyik típusa esetében a gyököcske eredetű gyökérág az egyed fejlődése során elpusztul vagy növekedése erősen lelassul, szerepét pedig a csíranövény sziklevelek magasságába eső szárrészéből, a mezokotilból kiinduló ún. mellékgyökerek veszik át (mellékgyökérzet vagy homorhiza). Hajtás eredetű gyökerek a sziklevelek feletti szárrészből is kialakulhatnak, többnyire a már kifejlett növényen. Ilyenkor beszélünk járulékos gyökérzetről. Ezt például rizómán, tarackon, legyökerező szárakon vagy dugványokon figyelhetjük meg. A legvékonyabb, felszívást végző gyökérágakat valamennyi gyökérrendszer esetében hajszálgyökereknek nevezzük.
A gyökérszerkezet alkalmazkodása a nedvességtartalomhoz
A hajszálgyökér zonációja és elsődleges szöveti szerkezete
A hajszálgyökerek hossztengelyük mentén mind szerkezetileg, mind pedig funkcionálisan elkülönülő zónákra tagolhatók (1. ábra). A merisztémákat tartalmazó régió az osztódási zóna, ahol az osztódó sejtek mitotikus aktivitásának köszönhetően a gyökér sejtjeinek nagy része kialakul. A zóna belsejében helyezkednek el az ősmerisztéma iniciálisai, amelyek között elkülöníthető egy alacsony mitotikus aktivitással rendelkező sejtcsoport, a néma centrum (centrális anyasejtek). Az ősmerisztémákból – a hisztogén elméletnek megfelelően – különböző szövettájakat kialakító elsődleges merisztémák jönnek itt létre. Kétszikűeknél a merisztematikus régió legkülső rétege a dermokaliptrogén, amelyből a gyökér bőrszövete (a rizodermisz) és a gyökérsüveg származik (+info). A promerisztematikus iniciálisoktól a gyökérnyak felé eső irányban találjuk a gyökér közbülső részén a sztélét kialakító plerómát, valamint az ezt hengerpalástszerűen körülvevő periblémát, amelyből az elsődleges kéreg szövetei jönnek létre.
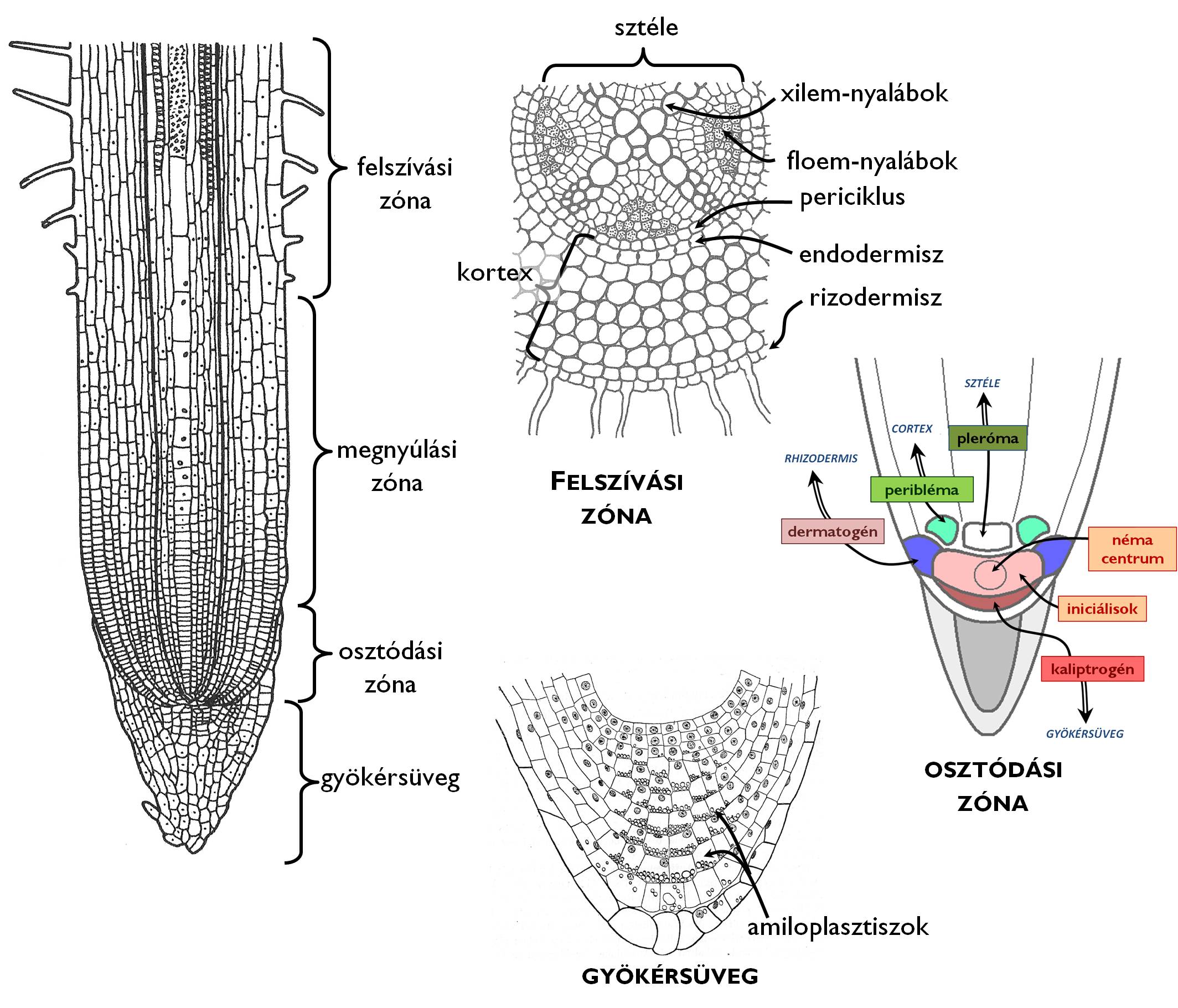
1. ábra. A gyökér zonációja.
(Jacob és mts. valamint Habelrandt ábrái nyomán)
Animáció a gyökér szerveződéséről: A hajszálgyökér zónáit bemutató ábrán a megfelelő zónákra állva megnézhető a megfelelő szakasz szerveződése.
A hajszálgyökér csúcsára sapkaszerűen borul rá a parenchimatikus sejtekből álló gyökérsüveg (kaliptra). Ennek legbelső régióját osztódóképességüket megtartó sejtek alkotják (összességük az ún. kolumella). A gyökérsüveg elsődleges feladata, hogy védje az osztódási zóna merisztémáit a sérülésektől (ami a gyökér növekedésének leállását okozná). A gyökérsüveg sejtjei különféle anyagokat termelnek és adnak le a környezetükbe, a talajjal érintkező sejtjei pedig elnyálkásodnak, leszakadoznak. Ez a két jelenség segíti a gyökér növekedését a talajban. A kaliptra amiloplasztiszokban gazdag sejtjei emellett a nehézségi erő irányának érzékelésében is részt vesznek, ami alapvető feltétele a gravitáció irányította növekedési mozgások (gravitropizmusok) kialakulásának.
Az osztódási zóna felett, a gyökérnyak felé haladva helyezkedik el a megnyúlási zóna. A hajszálgyökér sejtjei ebben a régióban intenzív vízfelvételüknek köszönhetően megnyúlnak. (Ebben a zónában a legintenzívebb gyökér hossznövekedése.) A megnyúlással párhuzamosan megindul a sejtek differenciálódása is: a zóna legfelső részén már kialakulnak az elsődleges szövetek.
A következő szakaszban megindul a megnyúlási zónában kialakult szövetek működése is; ez a felszívási zóna, amit zárvatermők esetében gyökérszőrös zónának is nevezünk. A régió nevét onnan kapta, hogy ez a zóna a tápanyagfelvétel színtere: itt lépnek be a talajból az ásványi anyagok és a víz a gyökér szöveteibe, illetve itt jutnak a szállító elemek belsejébe is, ahonnan aztán a megfelelő transzportfolyamatok eljuttatják azokat a növény egyéb részeibe. A néhány milliméter hosszúságú felszívási zóna felső régiójában a gyökérszőrök folyamatosan elhalnak, a rizodermisz leszakadozik, átlépünk a felszívást nem, csak transzportot végző szállítási zónába. Ezen a szakaszon jelennek meg a legtöbb gyökér esetében a másodlagos szövetek valamint az oldalgyökerek is. Emiatt nevezzük ezt a gyökérnyakhoz legközelebb eső szakaszt vastagodási ill. elágazási zónának is.
A felszívási zónából készült keresztmetszeti képen tanulmányozhatjuk a gyökér elsődleges szöveti felépítését. A bőrszövettel (itt: rizodermisz) borított szerv belső állománya – a száréhoz hasonlóan – két szövettájra különül: a középen elhelyezkedő, szállítónyalábokat tartalmazó központi hengerre (sztéle) és az ezt csőszerűen körülvevő elsődleges kéregre (kortex). A rizodermisz gyökérszőrein keresztül aktív transzporttal felvett ionok és az azokat passzívan (ozmózissal) követő víz először a kortex legkülső sejtsoraiba kerül. Abban az esetben, ha ez(ek) a réteg(ek) anatómiailag is elkülönülnek a kéreg egyéb sejtjeitől, exodermiszről beszélünk, amit elsősorban nedves vagy elöntött talajban levő gyökerekben találunk. Az exodermisz sejtjeinek falába paraanyag (szuberin) rakódik, ezzel egy védő funkciójú, vízzáró réteg jön létre, amelyen keresztül csak meghatározott helyeken juthatnak a tápanyagok a belsőbb régióba. A kortex fő állományát raktározó parenchima alkotja. A kortex legbelső, sztélével érintkező sejtsora az endodermisz. Az exodermiszhez hasonlóan az endodermisz sejtjeinek fala is parásodik, ezzel egy vízzáró réteget hozva létre a központi henger és a kéreg határán.
A vízzáró réteg változatossága: az endodermisz-típusok
A központi henger legkülső, endodermisszel határos sejtsora a merisztematikus aktivitásukat megőrzött sejtekből álló periciklus. E sejtrétegből indul meg az oldalgyökerek kialakulása. Az osztódóképes sejtek emellett részt vesznek a gyökeret vastagító oldalmerisztémák, a hullámos kambium és a fellogén kialakításában.
A sztéle fő állományát parenchimaszövet alkotja. Ebbe a szövetbe ágyazódva találjuk a gyökér egyszerű nyalábjait. A xylemnyalábok száma alapján megkülönböztetünk diarch, triarch, tetrarch pentarch stb. gyökereket. Általánosságban elmondható, hogy a Liliopsida [egyszikűek] osztályba tartozó fajok gyökere poliarch, azaz 8-10-nél több fanyalábot tartalmaz, és ezek a nyalábok nem érnek össze a gyökér középvonalában, a gyökérben tehát bélszövet található. A valódi kétszikűek (Rosopsida) gyökere ennél kevesebb xilem-nyalábot tartalmaz (oligarch gyökér), és ezek a fanyalábok gyakran összeérnek, ezzel egy csillag alakú, faelemekből álló struktúrát hozva létre a gyökér keresztmetszetének közepén.
A gyökér xilem-nyalábjainak jellegzetessége, hogy bennük a protoxilem elemek kifelé, míg a metaxilem elemek a középpont felé helyezkednek el (exarch xilem), mivel a xilem kívülről befelé kezd szerveződni. A háncsnyalábok a fanyalábok között, azokhoz képest excentrikusan helyezkednek el. Ezekben a protoelemek szintén a rhizodermisz felé, míg a metafloem tagjai a középvonal felé esnek (exarh floem). A gyökér szilárdító szöveti elemeit (elsősorban szklerenchima-rostokat) a fanyalábokhoz kapcsolódva találjuk, tehát a szerv szilárdítása annak hossztengelyére koncentrálódik. Így egy ún. kábelstruktúra alakul ki, amelynek előnye, hogy a lehető legkevesebb plusz anyag felhalmozásával ellenálló szerkezetet biztosít a növényt kihorgonyzó gyökérnek a húzóerőkkel szemben. (+info)
A gyökér másodlagos vastagodásának folyamata, a vastagodott gyökér felépítése
A Rosopsida osztályba [valódi kétszikűek] tartozó növényfajok gyökere szinte minden esetben másodlagosan vastagszik. (Ez a magyarázata annak, hogy miért a gyökérnyak közelében a legvastagabbak a gyökerek.) A Liliopsida [egyszikű] fajok gyökerein ez a folyamat nem figyelhető meg. Ebben az esetben a leszakadozó rizodermisz védő szerepét az exodermisz veszi át, miközben a kortex belső sejtrétege(i) másodlagos ill. harmadlagos endodermisszé alakulva egy belső záró réteget hoznak létre.
A másodlagosan vastagodó gyökerek esetében a felszívási zóna feletti régióban a gyökérszőrök eltűnésével párhuzamosan a xilem- és a floem-nyalábok közötti sztéle-parenchima dedifferenciálódni kezd és a sejtek visszanyerik osztódóképességüket. A fanyalábokon kívül és a háncsnyalábokon belül elhelyezkedő merisztematikus sejtek végül összekapcsolódnak egymással, és a periciklus fanyalábokkal szomszédos régióival együtt egy összefüggő gyűrűt, a jellegzetes lefutású hullámos kambiumot (gyökérvastagító kambiumot) hozzák létre (2. ábra). A hullámos kambium sejtjei – hasonlóan a vastagodó szárakban működő vaszkuláris kambiumhoz – periklinális osztódásaikkal kifelé másodlagos háncselemeket, míg befelé másodlagos faelemeket hoznak létre. A másodlagos szállítószövetek rétegződésének eredményeként a kezdetben valóban hullámos lefutású gyökérvastagító kambium egyre inkább kikerekedik, végül egy szabályos hengerpalást (keresztmetszetben kör) alakot vesz fel.
A másodlagosan vastagodott gyökér legbelső régiójában megfigyelhetők az elsődleges xilemnyalábok (oligarch gyökerek esetében csillag alakba rendeződve), ezen kívül a másodlagos faelemeket találjuk (ezek közül a legfiatalabb rétegek helyezkednek el kívül, a kambium közelében). A gyökér másodlagos fájának szerveződése a fatest felépítéséhez hasonló: évgyűrűs szerkezetet mutathat, benne a tracheák, tracheidák és farostok mellett bélsugarak futnak a kambium gyűrűjén kívül található másodlagos háncs irányába. A másodlagos háncselemek legfiatalabb képviselőit legbelül, míg idősebb rétegeit kívül, a kéreg maradványai alatt találjuk. Az elsődleges háncsnyalábokat a vastagodott gyökérben már nem találjuk meg, mivel annak vékony falú elemei összenyomódnak, eltűnnek (obliterálódnak).
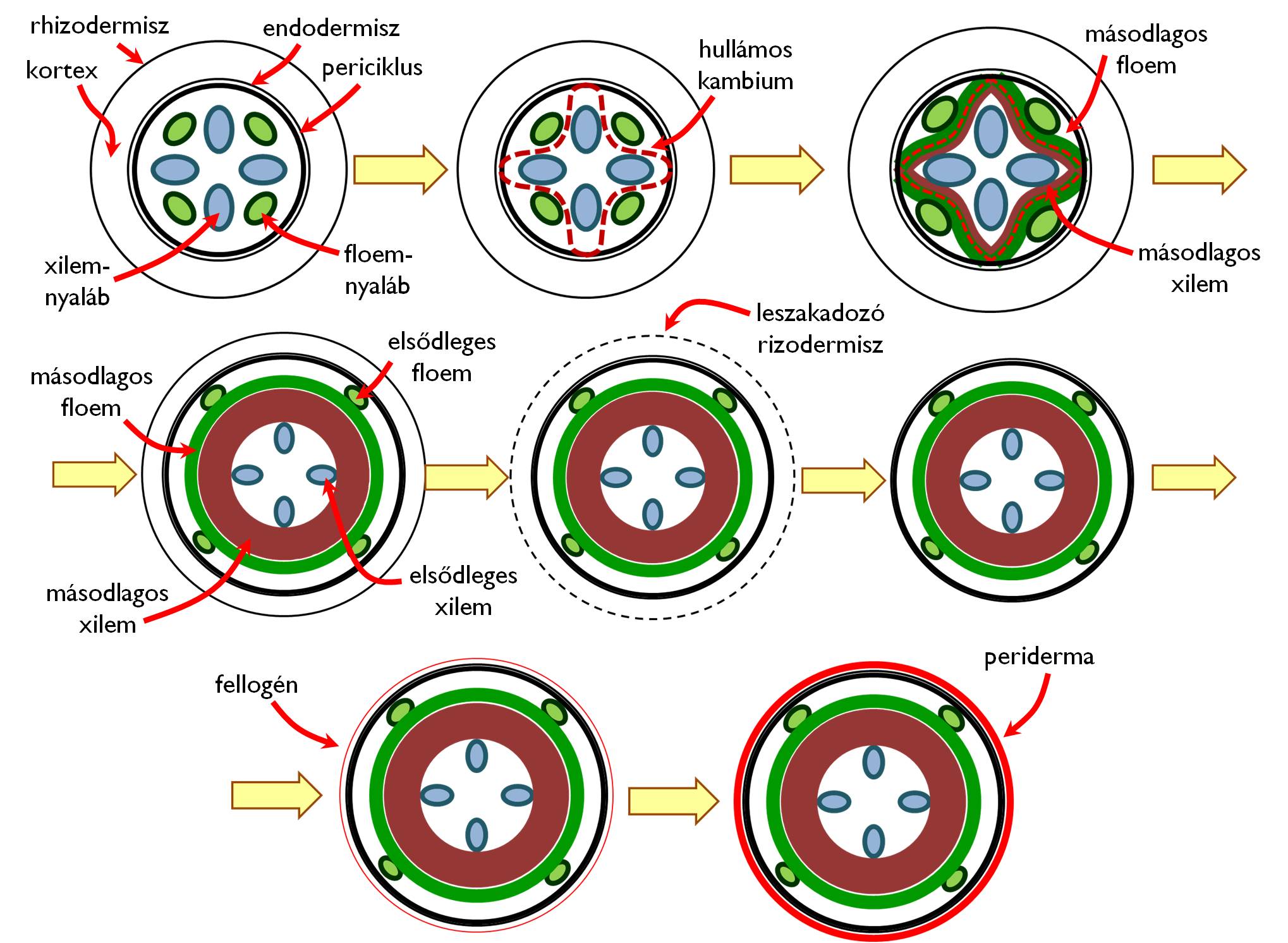
2. ábra. A gyökér másodlagos vastagodása.
A másodlagos szállítószövetek régióján kívül egy ideig még megtaláljuk az elsődleges kortex maradványát, ám mivel ennek sejtjei többnyire nem képesek követni a gyökérátmérő növekedését, ezért viszonylag korán leszakadozik. Helyén leggyakrabban másodlagos bőrszövet (periderma) jön létre. A peridermát kialakító fellogén (parakambium) a periciklusnak a hullámos kambium kialakításában részt nem vevő sejtjeiből jön létre, amelyek a fanyalábokkal szomszédos területeken (ahol a periciklus-sejtek a hullámos kambium részévé váltak) a kortex belső sejtrétegeiből másodlagosan létrejött osztódószöveti sejtekkel egészülnek ki összefüggő gyűrűvé (+info). A fellogén kifelé történő, egyirányú (monopleurikus) osztódása hozza létre a parásodott falú fellomot. Abban az esetben, ha a fellogén bipleurikusan működik, a periderma alatt egy megvastagodott fellodermából álló, elsősorban raktározó funkciójú másodlagos kéreg is létrejön.
Több kambiumgyűrű párhuzamos működése: polikambialitás a répatestben
A gyökér ökológiai alkalmazkodásának lehetőségei: a gyökérmódosulatok szövettana
A korábban említett alapfunkciók mellett a gyökér egyéb feladatokat is elláthat. Az így létrejövő módosult gyökerek esetében az új funkciók megvalósítása speciális szöveti alkalmazkodást igényel. A raktározó gyökerek (karógyökér, koloncos gyökérzet, ikergumó) esetében a fokozott tartalékoló funkció ellátása céljából fejlett raktározó parenchimából álló másodlagos kéreg jön létre a vastagodási zónában, amelynek sejtjeiben a különféle leukoplasztiszok (többnyire amiloplasztiszok) tartalmazzák az akkumulált tápanyagokat, amelyek segítségével a növény a következő vegetációs időszakban a hajtásait létrehozza majd (pl. Daucus carota subsp. sativus [sárgarépa], Dahlia × variabilis [dália]). Speciális raktározási módot jelentenek a szukkulens gyökerek (pl. Chlorophytum comosum [csokrosinda]), amikor a másodlagos kéregben víztartó alapszövet helyezkedik el.
A levegőben elhelyezkedő léggyökerek esetében lehetőség nyílik a fotoszintézisre is (zöld színű, ún. asszimiláló léggyökerek); ekkor a gyökér kérgi régiójának külső sejtsorai klorenchimává alakulnak át. A levegő páratartalmát hasznosító léggyökerek (pl. epifiton orchideák vagy a Monstera deliciosa [levélfa]) esetében a szövettani részben már említett, elhalt sejtekből álló, vízfelszívó képességű velamen radikumnak nevezett többrétegű bőrszövet borítja a gyökereket. A hajlító igénybevételnek kitett, támasztó funkciójú léggyökerek (koronagyökér, támasztó gyökér) specialitása a bőrszövet közelében elhelyezkedő rostkötegek megjelenése: ezekben a gyökerekben nagyméretű, kiterjedt bélszövetet tartalmazó központi hengert, és az ennek szélére tolódó, igen keskeny kéreg alatt elhelyezkedő nyalábokat és szklerenchima-rostokat találunk. A talajfelszínt áttörő légzőgyökerek (pneumatoforák) anatómiai sajátossága a kiterjedt aerenchima (átszellőztető alapszövet) jelenléte.
A más szervezetekkel együttélő növények gyökérmódosulatainak szövettani sajátosságai a kölcsönhatásban álló partnerek közti anyagátadást szolgálják. A szívógyökerek anatómiai felépítése alapvetően nem tér el a tápanyagok felszívására szolgáló gyökerekétől, a hajszálgyökerek azonban nem gyökérszőrök segítségével veszik át a vegyületeket a gazdától, hanem szívófonalakként (hausztórium) hatolnak a parazitált növény szállítószöveteibe (valódi paraziták esetében a háncsba, fotoszintézisre képes félparaziták esetében a farészbe).
A szimbiózisra szolgáló gyökerek esetében a szimbionta partner élhet a gazdanövény sejtjei közötti terekben, de akár a sejtek belsejében is. Számos növényfaj él tartósan együtt nitrogénfixációra képes mikroorganizmusokkal. A levegő nitrogénjét a növények számára is hasznosítható nitrogénformává (szerves nitrogénné vagy ammónium-ionná) prokarióta élőlények képesek alakítani. Kékalgák (Cyanobacteria) élnek például Cycadophyta törzs képviselőinek [szágópálmák] korallgyökereiben, egy összefüggő, kékes színű sejtréteget képezve a gyökér kortexében. A valódi baktériumok (Eubacteria) közé tartozó Frankia nemzetség képviselői hasonló réteget alkotnak az éger fajok (Alnus sp.) mikodomáciumnak nevezett gyökereinek kérgi régiójában. A nitrogéngyűjtő organizmusok és növények szimbiózisai közül a legismertebb a hüvelyesek (Fabaceae) gyökérgümőiben élő Rhizobium fajok és gazdanövényeik kapcsolata. Ezek a baktériumok a gyökérszőrökben kialakított infekciós csatornán át jutnak el a hajszálgyökerek kortexébe, ahol a növényi hormonokhoz hasonló hatású vegyületeket termelve sejtburjánzást váltanak ki. Az így létrejövő gümő belsejében szaporodnak el a baktériumsejtek, amelyek azonban nem a sejt közötti járatrendszerben, hanem a növényi sejtek belsejében élnek mint egyfajta speciális sejtalkotók (szimbioszóma). (+info)
A növényi gyökerek és gombafonalak együtteseként kialakuló, szimbionta eredetű szervmódosulat neve mikorrhiza. A növényfajok több mint 90%-a esetében megfigyelhető együttélésnek anatómiai alapon két fő kategóriáját különíthetjük el. Az ektomikorrhiza esetében a gombahifák egy összefüggő bevonatot (ún. gombaköpenyt) hoznak létre a hajszálgyökerek felületén, ahol így nem alakulnak ki gyökérszőrök. A köpenyből a gombafonalak behatolnak a gyökér bőrszöveti ill. kérgi sejtjei közé, ahol körülfonva azokat kialakítják az anyagátadásra szolgáló felületet. Ez a belső hifaszövedék a Hartig-háló. A gombaköpenyből a talaj irányába futó kiágazó hifák jelentősen megnövelik az anyagfelvevő felületet (ami a gyökérszőrök felületének is többszöröse). Ezen keresztül a gombapartner segíti a növény víz- és ásványianyag-felvételét, míg a növénypartner szerves anyagokat ad a gombának. (Ennek alapján már érthető, hogy miért létfontosságú az ektomikorrhiza a gyökérszőrökkel nem rendelkező nyitvatermő fajok számára.)
A mikorrhizák másik típusa az endomikorrhiza. Ebben az esetben a gombafonalak nem képeznek köpenyt a hajszálgyökér felületén, viszont behatolnak a kortikális (kérgi) sejtek belsejébe, ahol jellegzetes gombaképleteket hoznak létre. (Megjegyzendő, hogy a behatoláskor a hifák csak a sejtfalat törik át, a sejthártyát maguk előtt tolják; a növényi sejt tehát nem pusztul el.) Az endomikorrhizák legelterjedtebb típusa az ún.vezikuláris-arbuszkuláris mikorrhiza. Ekkor a gyökérsejtekben fácskaszerűen, gazdagon elágazó hifavégek ún. arbuszkulumok és hólyagszerű vezikulák jönnek létre. Az előbbiek képezik az anyagátadási felületet a szimbionta partnerek között, míg a vezikulák raktározásra szolgálnak. A talaj irányába az endomikorrhizából is gombafonalak futnak, amelyeknek a szerepe az ektomikorrhizák kiágazó hifáival egyezik meg.
A hajtás
A hajtás az embrió rügyecskének (plumula) nevezett pólusából kialakuló vegetatív szervek, a szár és a levél egysége. A valódi szövetes növények megjelenése során ez volt a legkorábban kialakult növényi szerv.
A hajtás evolúciója: a telóma-elmélet
A hajtás kialakulása
A hajtást alkotó szervek eredendően a csíranövény embrionális hajtáscsúcsából, a rügyecskéből (plumula) származtathatók. A több éven át fejlődő növények esetében, ha a vegetációs időszak nem folytonos, hanem a klimatikus viszonyok miatt azt nyugalmi időszakok szakítják meg, az aktív hajtáscsúcsok gyarapodása időről időre leáll és a kedvezőtlen időszakot ebben a formában vészelik át. Az embrionális állapotú, nyugalmi állapotban levő rövid szártagú hajtáskezdeményeket nevezzük rügyeknek. A rügyek elhelyezkedése a növényen – az ökológiai környezettől függően – többféle lehet. A mérsékelt övben ennek alapján különítjük el egymástól az ún. Raunkier-féle életformatípusokat (lásd a 6. fejezetben!).
A rügy részei: a nyugalmi állapotban levő hajtáscsúcs merisztémái, a következő évben kialakuló hajtástengely kezdeménye (rügytengely), az erre boruló levélkezdemények és többnyire az ezeket borító allevelek, a rügypikkelyek. A vegyes ill. virág(zati) rügyek esetében a generatív szervek kezdeményei is előfordulhatnak a rügyben.
A vegetációs időszak kezdetén a hajtáscsúcs működésbe lépésével megindul a hajtás növekedése. A hajszálgyökérhez hasonlóan a különböző fejlettségű régiók a hajtástengely esetében is megfelelő zonációban jelennek meg a hossztengely mentén (4. ábra). A hajtás végén találjuk az embrionális zónát, amely nem más, mint a hajtáscsúcsban található ős- ill. elsődleges merisztémák összessége.
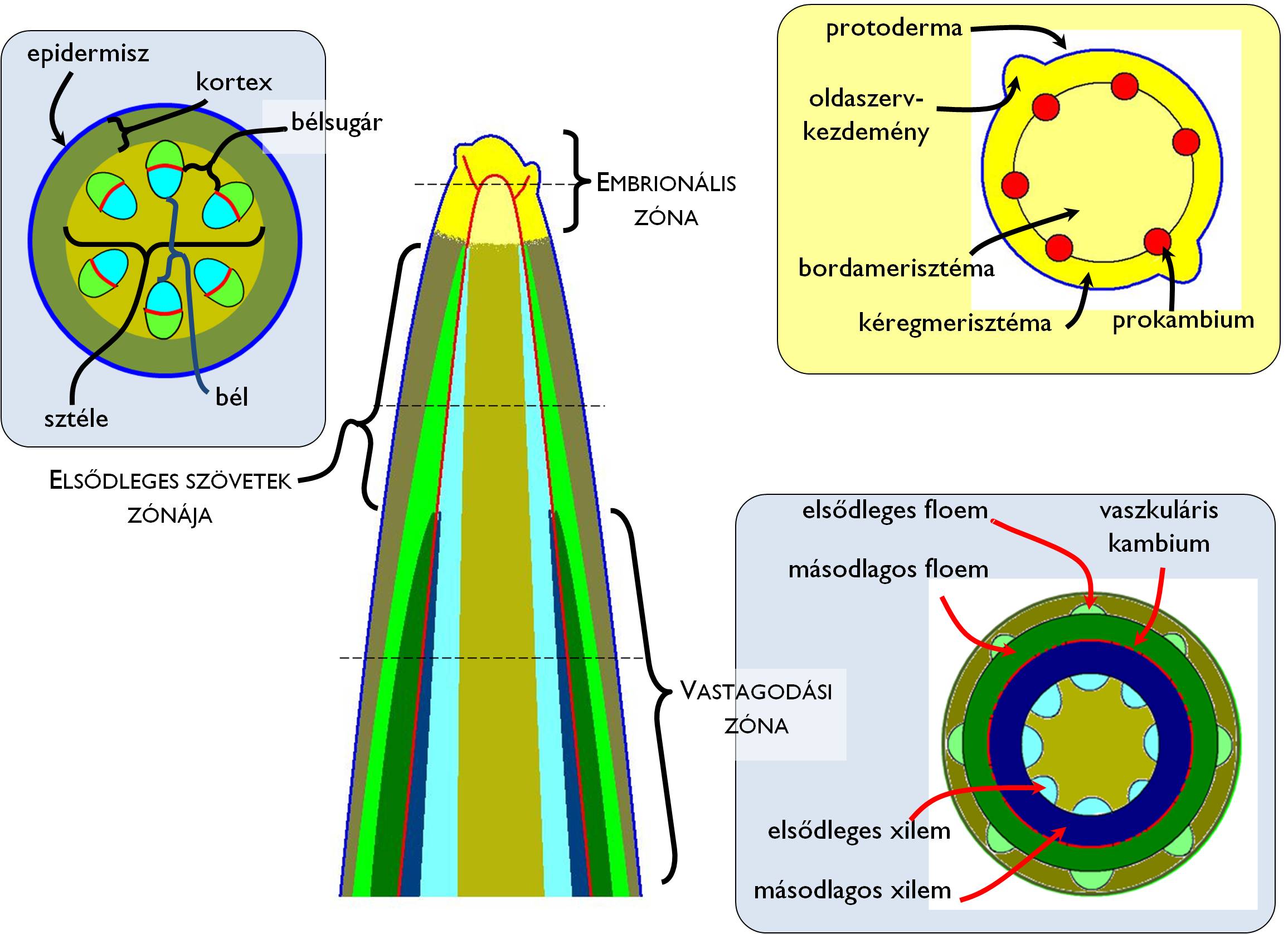
4. ábra. A hajtástengely zónái.
A szár szöveteinek kialakulása: a hajtáscsúcs merisztémái
Az osztódószövetek mitózisaival kialakuló sejtek a hajtáscsúcs alatti régióban megnyúlnak illetve ezzel párhuzamosan differenciálódnak (differenciálódási zóna) és kialakulnak a hajtástengely elsődleges szövetei. Ezt az elsődleges növekedési folyamatot a legtöbb szártípus (fás és lágy szárak esetében egyaránt) a másodlagos növekedés követi, ami kétféle módon valósulhat meg. Az elsődleges szövetek zónája alatt működésbe lépő másodlagos oldalmerisztémák (vaszkuláris kambium, parakambium) biztosítják a vastagodást. Emellett néhány lágyszár (szalmaszár, tőszár) esetében a szárcsomókban megmaradó merisztematikus sejtek aktivitása utólagos szártagmegnyúlást (interkaláris növekedés) eredményezhet.
A hajtástengely anatómiája
A hajtástengely elsődleges szöveti szerkezete
A hajtástengely (szár) elsődlegesen a növény tengelyéül szolgálva tartja az egyéb szerveket, részt vesz a szervek közti szállítási folyamatokban és – az esetek nagy részében – fiatalon fotoszintézist is végez. A külvilág felé – többnyire egyrétegű – epidermisz határolja, amelyen növényi szőröket, gázcserenyílásokat és kutikulát találunk. Belső szöveti állománya – a gyökérhez hasonlóan – két szövettájra, az elsődleges kéregre (primer kortex) és a központi hengerre (sztéle) osztható. Földfelszín feletti szárak esetében a kortex többnyire igen keskeny, csak néhány sejtrétegből áll.
A szár kortexében előforduló szövetek
A hajtástengely szállítószöveteket tartalmazó szövettája – a gyökérhez hasonlóan – a központi henger (sztéle), ami azonban nem válik el olyan élesen a kortextől, mint a gyökérben. Külső vonalát a szállítóelemek epidermisz felé eső felületének határvonala jelöli ki. Amennyiben a szár tengelyvonalában nincsenek szállítónyalábok, hanem alapszövet tölti ki, bélszövetről (ha itt üreg nyílik, bélüregről) beszélünk. A szállítóelemek között szinte minden esetben találunk sugárirányban rendeződő alapszöveti sejtsorokat; ezek a bélsugarak.
A szár központi hengerében – nyalábos sztéleszerkezet esetében – mindig összetett nyalábokat találunk. Kollaterális nyalábok esetében a xilem a szár középpontja, míg a floem a külvilág felé néz. A xilemben a protoxilem elemek befelé, míg a metaxilem elemek a háncsrész felé esnek (endarch xilem), míg a floem exarch szerveződésű, azaz a protoxilem elemek kifelé, a metaxilem tagjai a farész felőli oldalon helyezkednek el.
A hajtástengely másodlagos vastagodása
A hajtástengely elsődleges szöveti zónája alatt a legtöbb kétszikű szártípus esetében másodlagos (oldal)merisztémák jönnek létre, működésükkel pedig megindul a szár másodlagos vastagodása. A vastagodó szár másodlagos szállítószöveteit az összefüggő vaszkuláris kambium („kambiumgyűrű”) hozza létre, míg az átmérőbeli gyarapodás nyomán leszakadozó epidermisz szerepét átvevő peridermát a fellogén (parakambium) sejtjei termelik.
A nyalábos elrendeződésű elsődleges szállítószövetet tartalmazó szárak hajtáscsúcsában a prokambium is nyalábos elhelyezkedésű. A prokambium merisztematikus aktivitású leszármazottja a nyalábok belsejében is megmarad, ezért e szárakban kollaterális nyílt nyalábokat találunk. A másodlagos vastagodási folyamat megindulását megelőzően a nyalábokban levő kambium (nyalábkambium, faszcikuláris kambium) a nyalábok közti bélsugarak alapszöveti sejtjeinek dedifferenciálódásával kialakuló másodlagos merisztematikus aktivitású szakaszokkal (nyalábközti kambium, interfaszcikuláris kambium) kiegészülve hozza létre az összefüggő vaszkuláris kambiumot (+info).
A faszcikuláris kambium (néhány sejtsor vastagságú bélsugarak kivételével) mindig másodlagos szállítószövetet hoz létre, a nyalábközti kambium működésének terméke azonban változatos lehet (5. ábra). A látszólag összefüggő másodlagos szállítószövetben mindig bélsugarak helyezkednek el, ráadásul – a megfelelő klimatikus viszonyok között fejlődő, fás szárú fajok esetében – a fatest évgyűrűs mintázata is kialakul (l. később).
Az interfaszcikuláris kambium működésének sokfélesége: a szárvastagodás típusai
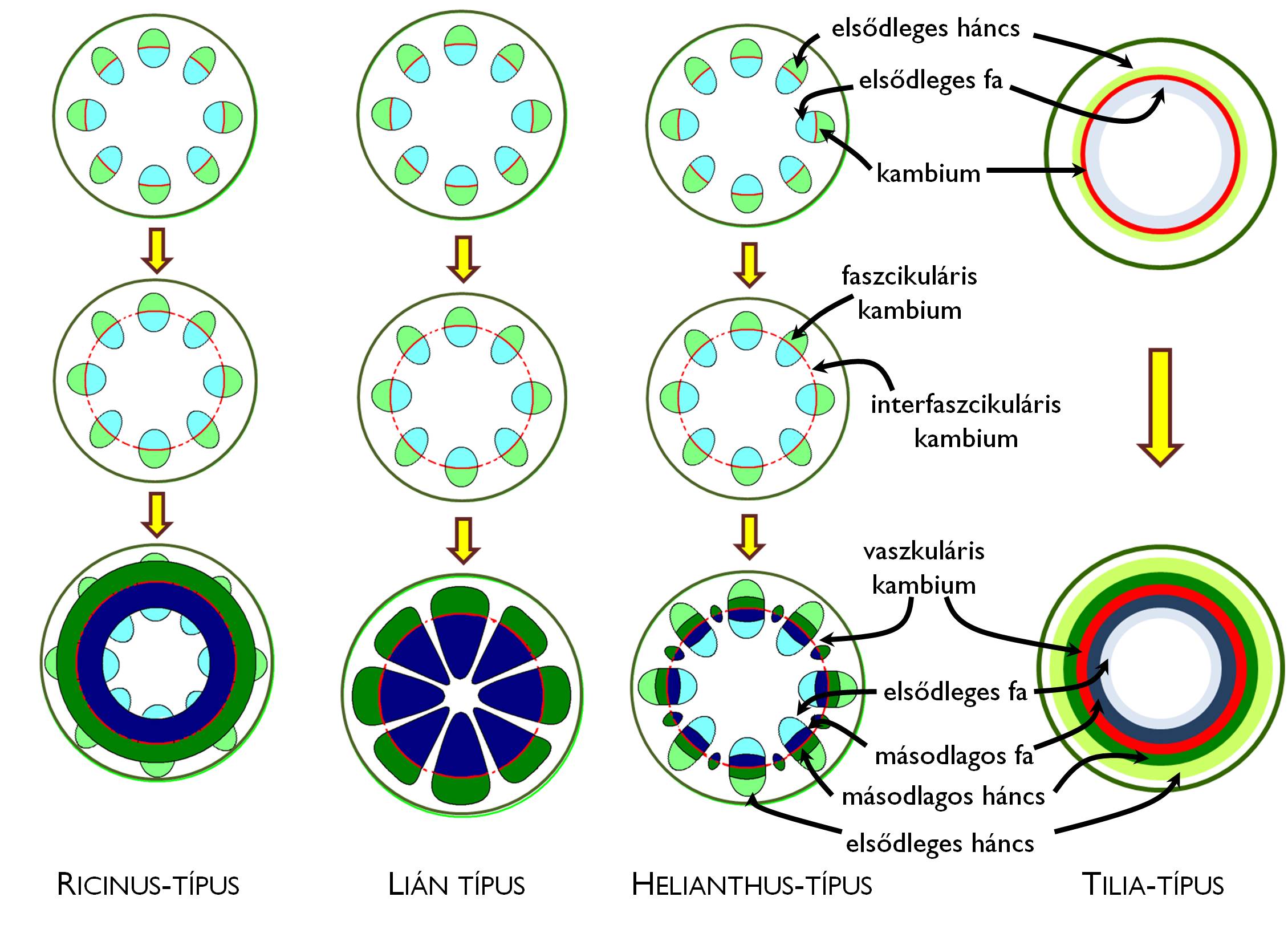
5. ábra. A szár másodlagos vastagodásának típusai.
A fás nyitva- és zárvatermő fajok szára a legtöbb esetben a nem nyalábos elsődleges szöveti szerkezetből kiinduló, ún. Tilia-típusú vastagodással fejlődik. A zöldhajtások hajtáscsúcsában e fajok esetében összefüggő prokambium-gyűrű található, így már az elsődleges fa- és háncselemek is összefüggő gyűrűt alkotnak (azaz nincsenek nyalábok). A kétféle szállítószövet között megmaradó, folytonos prokambium lényegében egy az egyben alakul vaszkuláris kambiummá a vastagodási zónában (+info), ahol megkezdi a másodlagos xilem és floem kialakítását. Ezzel jellegzetesen koncentrikus szöveti szerveződés jön létre: az elsődleges kortex alapszövete alatt az elsődleges floem gyűrűjét, majd a másodlagos floem (háncstest) rétegeit találjuk, ez után következik a kambiumgyűrű, majd a másodlagos xilem (fatest), ezen belül az elsődleges xilem gyűrűje, legbelül pedig a bélszövet parenchimája helyezkedik el.
A szárvastagodás speciális lehetősége: a fásodó szárú egyszikűek
A fás szár felépítése
A vastagodott fás szár fő tömegét a másodlagos szállítóelemek összessége, a háncstest és a fatest alkotja (6. ábra). A külvilág felé jellegzetes megalakulású, speciális másodlagos bőrszövet, a héjkéreg (ritidóma) határolja. Az alatta elhelyezkedő háncstest legkülső részén találjuk a legidősebb, míg a belső, a vaszkuláris kambium szomszédságában elhelyezkedő rétegeiben a legfiatalabb floem-elemeket. A kambiumgyűrű belső oldalán helyezkednek el a legfiatalabb, aktív szállítást végző faelemek, míg a szár közepe felé haladva egyre idősebb xilem-rétegek következnek. (Hangsúlyozandó tehát, hogy a szárban csak egyetlen kambiumgyűrű található!) A fa- és a háncstest között alapszöveti sejtek sugárirányba futó sorai, az ún. bélsugarak teremtenek kapcsolatot. Ezek az élő parenchima-sejtek a fás szárban folyó sugárirányú szállítás mellett tápanyag-raktározást is végeznek.
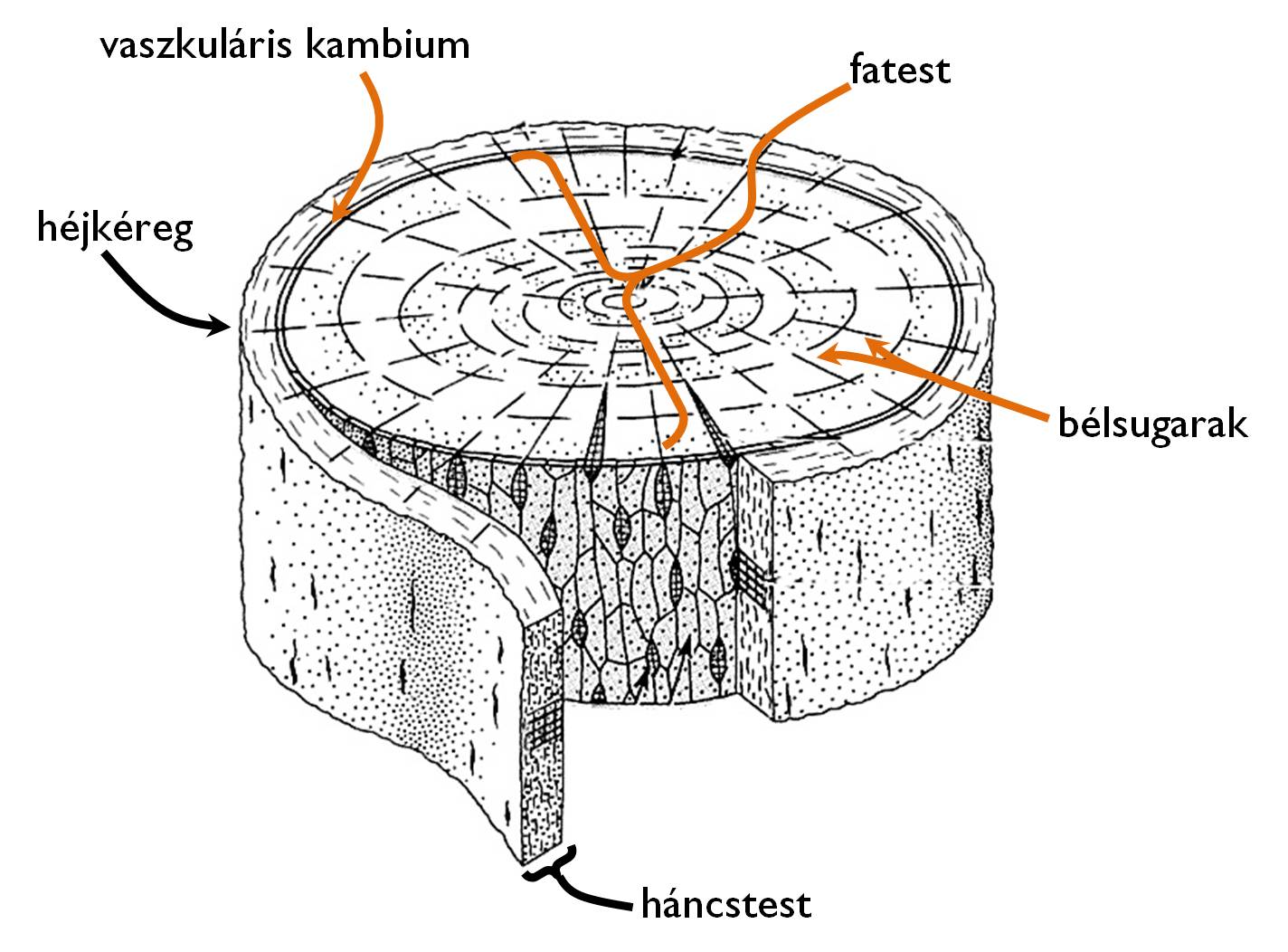
6. ábra. A fás szár szállítószövet-rendszereinek szerkezete. (Dános nyomán)
A fatest szerkezete
A fatest az évek folyamán belülről kifelé (centrifugálisan) növekszik, a kambiumgyűrű mind újabb rétegekkel gyarapítja állományát. Ennek megfelelően a legfiatalabb, a víz- és ásványianyag szállításában aktívan részt vevő faelemeket a fatest külső felületén találjuk; e rétegek összessége a szíjács. Az idősebb, a szár középvonala felé elhelyezkedő xilemsejtek már nem vesznek részt a szállításban, az itt elhelyezkedő tracheákat, tracheidákat töltősejtek (tilliszek) tömik el. A sejtek falában a fakorhasztó mikróbák (elsősorban gombák) szaporodásának megakadályozása céljából speciális másodlagos anyagcseretermékek (polifenolok) rakódnak. Ez a belső régió a geszt. Egyes fafajok (pl. Fagus [bükk], Quercus [tölgy], Pinus [fenyő]) – az ún. gesztesedő fák – esetében az anyagberakódás olyan mértékű, hogy a kivágott fa vágási felületén (bütüjén) is egyértelműen elkülönül a geszt állománya (+info).
A fatestet – a farészhez hasonlóan – elsősorban a különféle vazális elemek, parenchimasejtek valamint farostok alkotják. A nyitvatermők fájában csak tracheidákat, parenchimasejteket és rosttracheidákat találunk (farostokat és tág üregű tracheakat nem), ezért ezen fajok fateste szöveti szerkezetének képe homogén (homoxil fák). A zárvatermők farészének összetétele sokkal változatosabb (tracheidákat, tracheakat, farostokat, faparenchimasejteket és egyes fajok esetében rosttracheidákat is tartalmaz), anatómiai felépítése tehát igen heterogén – ezek az ún. heteroxil fák.
A fatestben a parenchimasejtek két rendszert alkotnak: a bélsugarak radiálisan rendeződött sorait, valamint a vazális elemeket tengelyirányban kísérő (szűk értelemben vett) faparenchima-sejtek rendszerét.
A xilem-elemek sejtfalának lignifikáltsága alapján puha- és keményfákat különböztetünk meg egymástól. Az alacsonyabb (40% vagy annál alacsonyabb) lignintartalmú puhafák gyors növekedésű, törékeny (töréskárokra, széldöntésre érzékenyebb) fajok (pl. Salix [fűz], Populus [nyár], Pinus [fenyő] fajok, Acer negundo [zöldjuhar]), amelyeket első sorban a papíripar hasznosít. Ezzel szemben a több lignint tartalmazó keményfák igen ellenállóak, a bútor- és építőipar számára értékesebb faanyagokat szolgáltatnak, biztonságosabban ültethető parkfák, útsorfák (pl. Fagus [bükk], Quercus [tölgy], Taxus [tiszafa], Prunus avium [cseresznye]). (+info)
Egyes éghajlati területeken a változó klimatikus körülmények miatt nem folyamatos a növények növekedése, azt nyugalmi periódusok szakítják meg. A mérsékelt égövben pl. az évszakoknak megfelelően működik a vaszkuláris kambium és ennek következtében jellegzetes, évgyűrűs mintázat alakul ki a fatestben (+info). Tavasszal, a fák rügyfakadását követően a nagy mennyiségű csapadéknak köszönhetően a növények rövid idő alatt nagy mennyiségű vizet vesznek fel és szállítanak a lombozat irányába. Ekkor tágabb üregű, vékonyabb falú szállító elemek (tracheidák, tracheák) keletkeznek, míg a rostok (ill. rosttracheidák) aránya ilyenkor alacsonyabb. Az ebben az időszakban kialakuló, makroszkóposan világosabb színű xilem-sávot nevezzük korai (tavaszi) pásztának 7. ábra). Nyár második felében illetve az őszi időszakban ezzel szemben lecsökken a szár folyadékszállításának üteme, szűkebb üregű, vastagabb falú vazális elemek jönnek létre és nagyobb arányban keletkeznek rostok, rosttracheidák. Így jön létre a sötétebb színű kései (őszi) pászta. A téli időszakban leálló – vagy lelassuló – kambiumműködést követően tavasszal hirtelen meginduló osztódási aktivitásnak köszönhetően az előző évi kései és az azévi korai pászta között egy éles (látszólagos) határvonal jön létre, ez az évgyűrűhatár. A fatest (azaz a fásodott szárrész) korát tehát a bütün látható világos VAGY sötét gyűrűk leszámlálásával határozhatjuk meg.
Az évgyűrűk vastagságát az adott vegetációs időszak időjárási illetve egyéb környezeti viszonyai határozzák meg: csapadékos években, ha elegendő tápanyag áll a növény rendelkezésére vastagabb, míg kedvezőtlen időszakban, esetleg egy súlyosabb kártevőhatás következtében keskenyebb évgyűrű (vagy pászta) jön létre. Az évgyűrűmintázat tehát lenyomatként őrzi a fát ért ökológiai hatások történetét. (+info)
A heteroxil fák évgyűrűiben a tracheák többféleképpen helyezkedhetnek el (7. ábra). A szórt likacsú fák esetében a nagyobb méretű tracheák a korai és a kései pásztában egyaránt előfordulnak (pl. Acer [juhar], Aesculus [bokrétafa], Fagus [bükk], Magnolia [liliomfa], Tilia [hárs]). A gyűrűs likacsú fák fatestében azonban a tág üregű tracheák a korai pásztában csoportosulnak (pl. Fraxinus [kőris], Catalpa [szivarfa], Morus [eperfa], Quercus [tölgy], Robinia [akác], Ulmus [szil]).
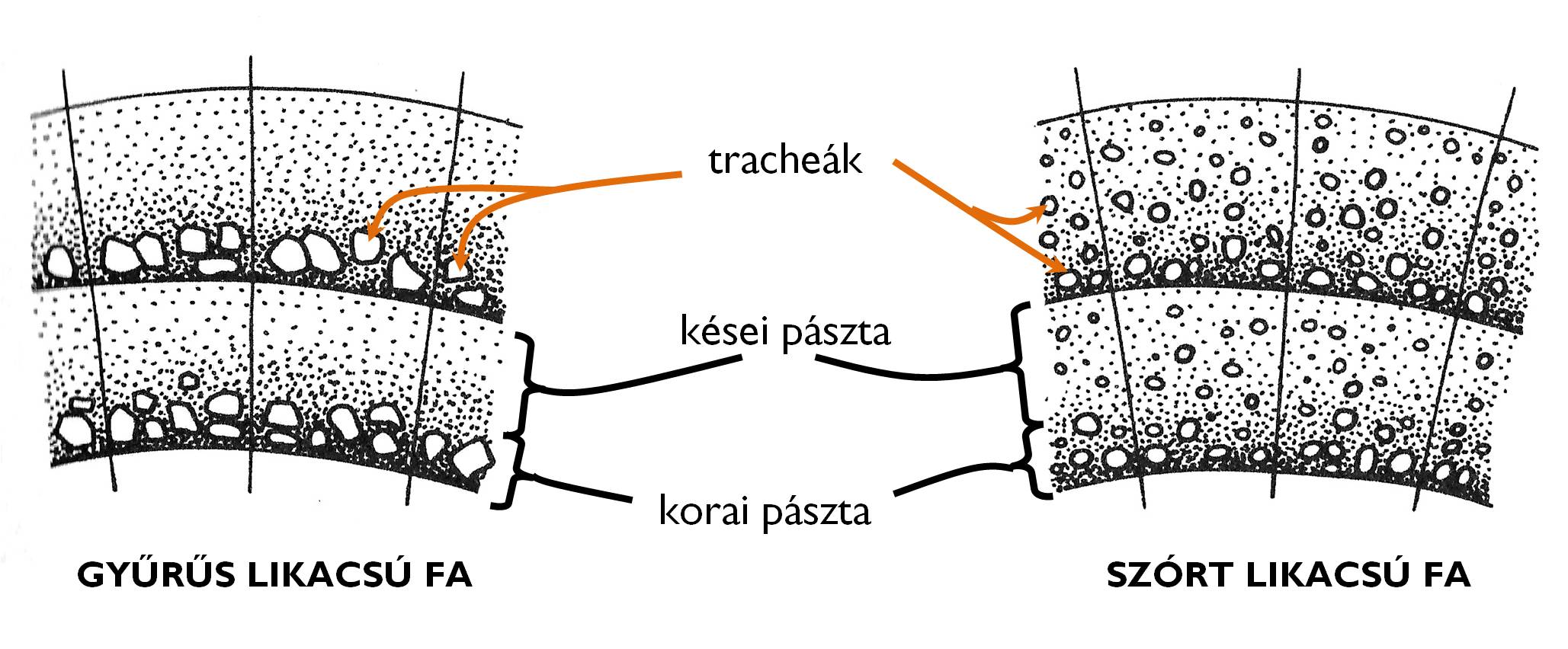
7. ábra. A gyűrűs és a szórt likacsú fa fatestének szerkezete.
(Jacob és mts. nyomán)
A háncstest szerkezete
A háncstest esetében is igaz, hogy legfiatalabb részei a vaszkuláris kambiummal határosak, ebben az esetben azonban ez azt jelenti, hogy ezek a szár közepe felé néznek, míg az idősebb, szállítást már nem végző rétegek a héjkéreg felőli oldalon vannak. A másodlagos háncs kialakulása tehát centripetális.
A fatesthez hasonlóan a háncstest is réteges szerveződésű. A háncsrostokban gazdag kemény háncs és a szállítóelemek túlsúlyával jellemezhető lágy háncs rétegei váltakoznak egymással. Ezek kialakulásának ritmusa azonban nem feltétlenül követi az éves periódusokat, így ezek számából nem következtethetünk a szár korára. A bélsugarak természetesen a háncstestben is folytatódnak, az átmérőbeli gyarapodás miatt azonban ezek a szár kerülete felé haladva kitágulnak, keresztmetszetben tölcsérszerűen tűnnek elő (bélsugár-tölcsérek).
A héjkéreg (ritidóma) kialakulása
A szár vastagodását az epidermisz – hasonlóan egyéb vastagodó szervek bőrszövetéhez – nem képes követni. A leszakadozó epidermisz helyét a másodlagos bőrszövet (periderma) veszi át. Az ezt kialakító fellogén az epidermisz vagy az alatta levő alapszöveti sejtek dedifferenciálódásával jön létre (+info). Már a beérett azévi hajtásokat is a fajra jellemző mintázottságú és morfológiájú másodlagos bőrszövet borítja.
A hajtás további vastagodásával a periderma is felrepedezik, leszakadozik. Ekkor a bőrszövet alatt elhelyezkedő kortex további rétegei alakulnak fellogénné, így továbbra is szabályos másodlagos bőrszövet borítja a szárat. Egyes fajok esetében (pl. Armeniaca [kajszi], Malus [alma]) még az idősebb szárrészeken (gallyakon, ágakon) is peridermát találunk. Egy idő után azonban elfogynak a kéreg alapszöveti sejtsorai, és a periderma a háncstesttel érintkező helyzetbe kerül. Ekkor a parakambium már a háncstest külső részének alapszöveti elemeiből dedifferenciálódik, osztódásaival pedig olyan bőrszöveti rétegek jönnek létre, amelyekben elhalt háncselemek (rostacsövek, rostasejtek, háncsrostok) maradványai is megfigyelhetők. Ezt a módosult felépítésű, háncselemeket is tartalmazó, vastag védőszövetet nevezzük ritidómának (héjkéregnek). A héjkéreg a szár további tágulása miatt szintén szétfeszül, aminek köszönhetően a fajra jellemző módon felrepedezik, lehámlik vagy lefoszlik. Eközben a háncstestnek újabb és újabb rétegei alakítják ki a fellogént. (Ez is magyarázza, hogy miért nem következtethetünk a háncstest vastagságából vagy rétegeinek számából a fa korára.)
A héjkéregben – rendszertani csoporttól függően – kristályok vagy különféle másodlagos anyagcseretermékek, színanyagok rakódhatnak le. Ez adja a különböző fajok ritidómájának jellegzetes megjelenését, állagát. A Quercus suber [paratölgy] esetében például a héjkéreg sejtjeinek fala nagy mennyiségű szuberint tartalmaz. Ez a leválasztott héjkéreg a „parafa”.
A levél
A levél evolúciós eredete: a mikrofillum és a makrofillum
A levél párhuzamos evolúcióval több törzsfejlődési vonalon is kialakult. Legkorábban a különböző ősharasztoknál jelent meg. Az egyes rendszertani csoportok fotoszintézist végző lombleveleinek anatómiája azonban eltérő lehet. Alapvetően két fő típus különíthető el, a mikrofillum és a makrofillum (8. ábra). A Lycopodiophyta törzs [korpafűfélék] képviselőire valamint a Monilophyta törzs Equisetopsida osztályára [zsurlók] jellemző mikrofillum esetében a levél szállítónyalábjai a szár nyalábjainak egyszerű oldalágaiként kanyarodnak ki a levélnyélbe. A mikrofillum tehát valójában nem más, mint a hajtástengely egyféle felszíni függeléke (emergenciája). A Monilophyta törzs páfrányokat tartalmazó osztályaiban kialakuló makrofillum szállítószövete teljesen más módon kapcsolódik a szár sztéléjének szállítószöveteihez. A hajtástengely szállítószövet-hengeréből kiválik egy (vagy több) nyaláb (levélnyomnyaláb) és a makrofillum levélnyelébe kanyarodva fut a levélbe. Ennek eredményeképpen azonban a szár szállítószöveteiben egy hiátus marad, az ún. levélrés. A levélnyomnyalábok kikanyarodása miatt a korai harasztoknál még hengerszerűen összefüggő szállítószövetrendszer (protosztéle, szifonosztéle - l. korábban!) feltagolódik és nyalábossá (eusztéle, diktiosztéle) válik. Az evolúció során tehát a makrofillum kialakulása okozta a nyalábos sztéleszerveződés megjelenését. Természetesen a levényomnyaláb kikanyarodásával nem szűnik meg a hossztengely mentén történő szállítás a szár adott szakaszán. A szállítószövetek oldalirányban kitérve, a levélrés felett ismét összezárulnak, tehát a szállítónyalábok rendszere nem egyenes csövekként, hanem oldalirányban anasztomizáló hálózatként képzelendő el. A páfrányok képviselői mellett valamennyi nyitva- és a zárvatermő faj levele is természetesen makrofillum.
A mikro- és makrofillum evolúciós eredete
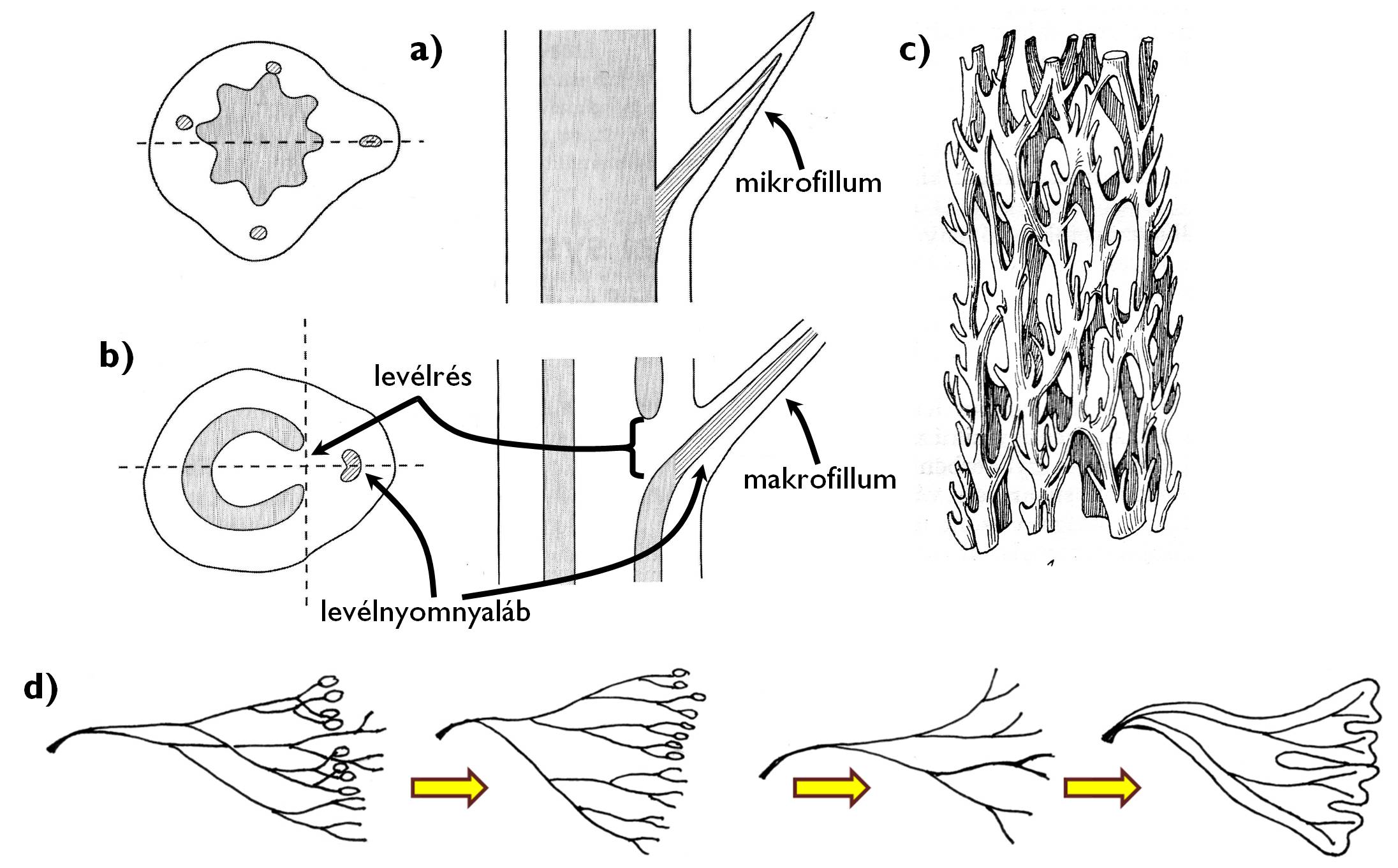
8. ábra. A mikrofillum és a makrofillum.
a) a mikrofillum kapcsolata a szárral; b) a makrofillum kapcsolata a szárral; c) a makrofillumos növények hajtásaiban megfigyelhető nyalábrendszer; d) a makrofillum evolúciós eredete a telóma-elmélet szerint.
(a-b: Beck nyomán, c: Kárpáti nyomán, d: Podani nyomán)
A levél kialakulása
A hajtás fejlődésénél már leírtuk, hogy az oldalszervek kezdeményei – így a levélprimordiumok is – már a hajtáscsúcs elsődleges merisztematikus régiójában megjelennek. Habár egy ideig megőrzik radiális szimmetriájukat, elég hamar ellaposodnak és növekedésük korlátolttá válik.
A lomblevelet kialakító merisztémák
A levél növekedésének és differenciálódásának általános szabályszerűségei rendszertani csoportonként eltérőek lehetnek. A páfrányok makrofillumainak növekedése például egészen sokáig korlátlan marad: miközben a levél alaphoz közeli részén már a lemez is teljesen kialakult, a levél csúcsa még differenciálatlan osztódószövetet tartalmaz, ami folyamatosan gyarapítja a levél állományát. A csúcsi rész sérülése esetében természetesen nem alakulna ki a teljes levél, éppen ezért ez a merisztematikus régió védett helyen, a pásztorbot-szerűen feltekeredett levél már kialakult szöveteinek védelmében helyezkedik el.
Hasonlóan korlátlan marad a levéllemez sejtjeinek gyarapodása a legtöbb pázsitfűféle (Poaceae) esetében. Ekkor azonban nem a levél csúcsán, hanem a levéllemez és a levélhüvely érintkezési vonalában marad egy merisztematikus zóna, ami folyamatosan pótolja a levéllemezt. Ez a magyarázata annak, hogy miért növekszik folyamatosan a legtöbb pázsitfűféle lelegelt vagy levágott levéllemeze. (+info)
A fás szárú növények leveleinek egy része a rügyek belsejében lényegében embrionális állapotban vészeli át a vegetációs időszakok közti nyugalmi periódust. Megjegyzendő azonban, hogy a rügydifferenciáció időszakára a levélszövetek szinte valamennyi sejtje kialakul, így a rügypattanást követően már nincs szükség további osztódásokra. Így a rügyek fakadása után már csak a vízfelvétel okozta sejtnövekedés biztosítja a levelek kiterülését, ami lényegesen gyorsabb folyamat, mint az osztódáson alapuló növekedés.
A levél szöveti szerkezete
A levél szárral érintkező része a levélalap. Itt kanyarodnak ki a hajtástengelyből a levélnyomnyalábok a levélnyélbe (ülő levelek esetében közvetlenül a levéllemezbe). (+info) A levélnyélben egyszikű fajok (Liliopsida) esetében több, valódi kétszikű növények (Rosopsida) esetében többnyire csak egy vagy néhány nyomnyaláb fut, alapszövetbe ágyazva. A nyél anatómiája valamelyest a száréhoz hasonló, azzal a különbséggel, hogy benne valamennyi szállítónyaláb egy irányba rendeződik: a farész a szár irányába eső oldal, azaz a színi (adaxiális) oldal felé, míg a nyalábok háncs része a fonáki (abaxiális) oldal felé néz. A levélnyelet minden esetben szilárdító alapszöveti elemek (a nyalábokat kísérő szklerenchimarostok, valamint a bőrszövet alatt elhelyezkedő kollenchima rétegek) merevítik. Zöld levélnyelek esetében természetesen az epidermisz alatt klorenchima is található.
A levéllemezbe lépő erek a levélerezet típusának megfelelő rendszerben oldalágakat képeznek, és a legvékonyabb szállítónyalábok révén az egész lemezt behálózzák. Természetesen nem minden levélsejthez fut külön szállítónyaláb, a vakon végződő nyalábok a lemeznek mindig egy-egy kicsiny részét látják el tápanyagokkal (illetve szállítják el innen a szerves anyagokat). Ezek az egységek az ún. érszigetek.
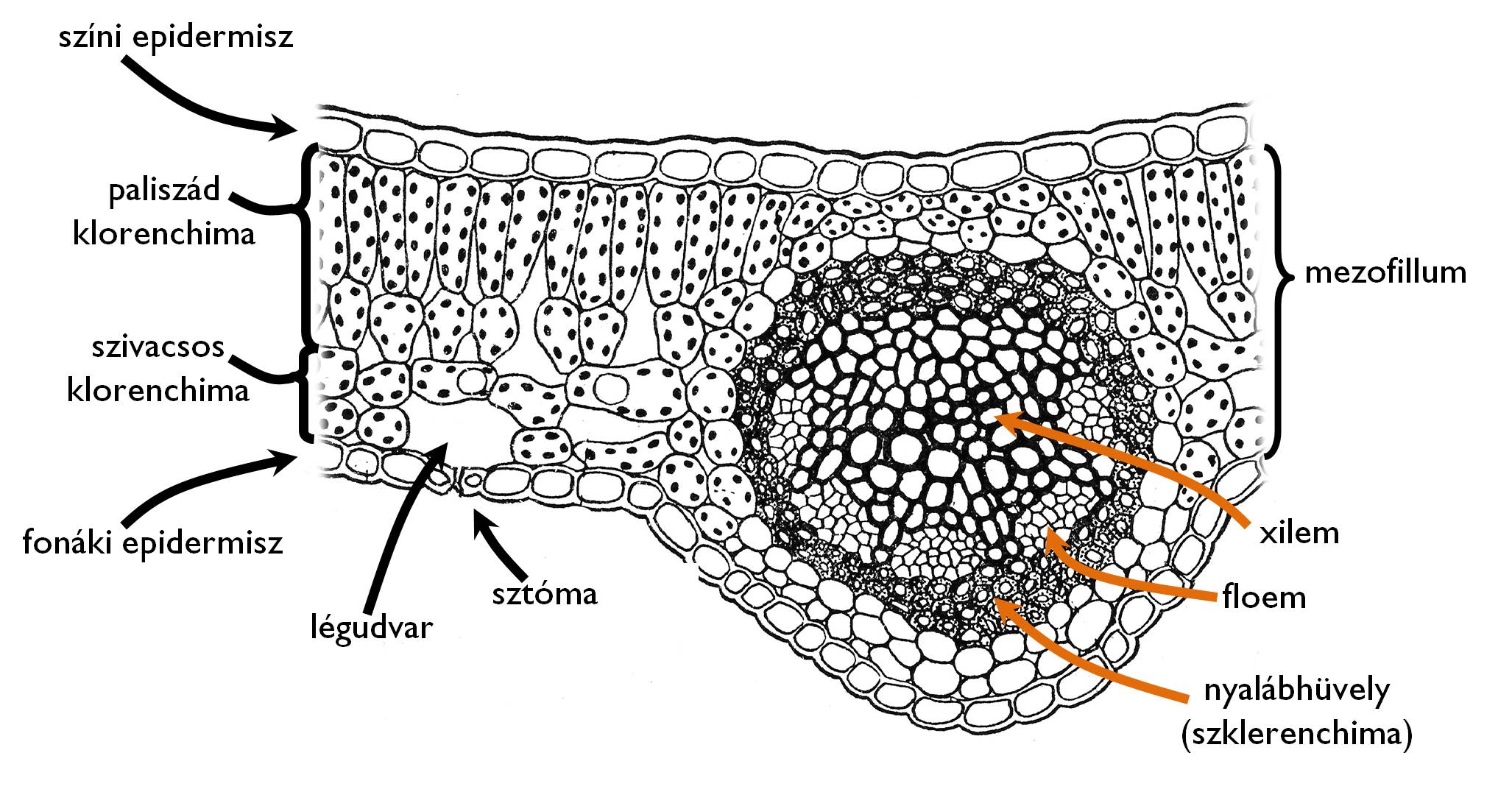
9. ábra. A mezomorf dorziventrális, bifaciális levél szöveti szerkezete.
(Sárkány és Szalai nyomán)
A levél keresztmetszeti képén a színi és a fonáki epidermisz között találjuk a levél fő szöveti állományát kitevő mezofillumot (9. ábra). Az átlagos ökológiai körülmények között kialakuló (mezomorf) levelek esetében a levél sztómáinak többsége a fonáki epidermiszen helyezkedik el (hiposztomatikus levél). A mezofillum nagy részét klorenchima alkotja, amely kétféle szövettípusból áll. A színi oldal felé a lemez felületére merőlegesen megnyúlt, szorosan illeszkedő oszlopos fotoszintetizáló sejtek helyezkednek el – ez a paliszád klorenchima rétege. A fonáki oldalon a szabálytalan alakú klorenchima-sejtek között intercelluláris üregekből álló skizogén járatrendszer biztosítja a sejtek gázcseréjét (szivacsos klorenchima). A sejtközötti járatrendszer külvilágba vezető része a gázcserenyílás légrése alatt elhelyezkedő légudvar. A kétféle klorenchima között átmenetet képeznek a gyűjtősejtek. Ezek olyan elágazó, oszlopszerűen megnyúlt sejtek, amelyek a színi oldal felé több paliszád sejttel, a fonáki oldal irányában pedig egy-egy szivacsos klorenchima-sejttel kapcsolódnak. A levél keresztmetszetén természetesen megtaláljuk a szállítónyalábokat (azaz a levélereket). Ezek a nyalábok kollaterális (esetenként bikollaterális) összetett nyalábok, xilem részük – a levélnyélhez hasonlóan – a színi, míg floemjük a fonáki oldal felé néz. Korlátolt növekedésű szerv lévén a levél zárt nyalábokat tartalmaz, habár az örökzöld, hosszabb ideig fejlődő levelek nyalábjaiban a növekedési időszak végéig megmarad a kambium (nyílt nyalábok). A levelet merevítő szilárdító elemek zöme a lemezt gerendázatszerűen kifeszítő erezet környékén helyezkedik el, a nyalábokat kísérő szklerenchimarostok, valamint hipodermálisan elhelyezkedő kollenchimarétegek formájában.
A hajtás többi részéhez hasonlóan természetesen egyéb szöveti elemek is előfordulhatnak a levelekben. Kiválasztást végző sejtek fordulhatnak elő az epidermiszben mirigyszőrök formájában, vagy akár a mezofillumban is. A tejnedves családok (Euphorbiaceae [kutyatejfélék], Apocynaceae [meténgfélék], Campanulaceae [harangvirágfélék], Papaveraceae [mákfélék] stb.) leveleiben tejcsöveket, a gyantás nyitvatermők leveleiben gyantajáratokat találunk. Néhány faj (pl. Laurus nobilis [babér], Hypericum perforatum [közönséges orbáncfű]) levélszöveteiben illóolajtartók fordulnak elő. Gyakran kristályzárványokat tartalmazó idioblasztok is megfigyelhetők akár az epidermiszben (pl. a Ficus [füge, fikusz] fajok esetében) akár az alapszövetben (pl. Malus [alma]). A fokozott szilárdságot igénylő levelekben (pl. vízfelszínen úszó, ún. natansz levelek esetében – lásd később!) a mezofillum elszórtan szklereidákat tartalmazhat.
A lomblevél szövettani típusai
A lombleveleket két szövettájuk, az epidermisz és a mezofillum szövettani felépítése alapján csoportosíthatjuk (10. ábra). Mindkét esetben a színi (adaxiális) és fonáki (abaxiális) oldalon levő szövetek anatómiai viszonya szolgál a csoportosítás alapjául. A két oldal megállapításához a szállítónyalábok orientációja nyújt segítséget: a farész a színi, a háncsrész a fonáki oldal felé mutat.
Csoportosítás a mezofillum szerkezete alapján
A levelek többségének mezofillumában a klorenchima szerveződése az előző részben bemutatott leírásnak megfelelő (a színi oldal felé paliszád, míg a fonáki oldal felé a szivacsos klorenchima néz). Ezeket az ún. dorziventrális leveleket színi oldaluk felől éri a fény, ezért az adaxiális oldalon a jobb fénykihasználó tulajdonságú paliszád réteget találjuk. Abban az esetben, ha a levél szorosan a szárra simul (pl. Thuja [tuja], Platycladus [életfa] vagy pikkelylevelű Juniperus [boróka] fajok esetében), akkor a fonáki oldal felől érkezik a mezofillumba a fény, és az előbbi szerkezet megfordul: az oszlopos réteg a fonáki, a szivacsos réteg a színi oldal felé áll (inverz dorziventrális levél). Az előző két levéltípus közös jellemzője, hogy a színi és a fonáki epidermisz alatt különböző szerkezetű alapszövetet találunk (heterolaterális levelek). Az izolaterális levél esetében a két epidermisz alatt azonos szerkezetű a klorenchima. Ezek a levelek többnyire mindkét oldaluk felől azonos mennyiségű fényt kapnak. Az izolaterális szerkezet nem feltétlenül jelenti azt, hogy csak egyféle felépítésű asszimiláló szövetet tartalmaz a levél. A heterogén izolaterális levél epidermiszei alatt oszlopos, míg közbülső állományában szivacsos (vagy legalábbis izodiametrikus sejtekből álló) klorenchimát találunk. A homogén izolaterális levélben azonban csak egyféle, többnyire kevés intercellulárissal átjárt, kerekded sejtekből álló klorenchimát találunk (pl. az Iris [nőszirom] vagy Viscum [fagyöngy] fajok levelében).
Csoportosítás az epidermisz szerkezete alapján
Abban az esetben, ha a levél színi és fonáki bőrszövete eltérő (elsősorban a sztómák eltérő gyakorisága miatt), bifaciális levélről beszélünk. Ez a leggyakoribb szövettani típus, amit elsősorban a párologtatás minimalizálása és a fény lehető legjobb kihasználása alakít ki. Ekvifaciális az a levél, amelynek színi és fonáki oldalát azonos felépítésű bőrszövet borítja. Ez a szerkezet akkor alakul ki, ha a levéllemez mindkét oldalát azonos intenzitással világítja meg a nap, ezért nem jelenti a vízvesztés (ill. a felmelegedés) minimalizálását a bőrszöveti képletek egyenlőtlen elosztása. Ekvifaciális levele van például a Viscum [fagyöngy], Loranthus [fakín] vagy a Pinus [fenyő] fajoknak. Speciális helyzet áll fenn az unifaciális levelek esetében. Ezen növények esetében az egységes epidermisz-szerkezet hátterében az áll, hogy az egyedfejlődés során eltűnik a levél színi oldala, ezért a levéllemezt minden oldalról (a szükségszerűen azonos) fonáki epidermisz borítja. Unifaciális levél kialakulásához kétféle fejlődési folyamat vezethet (10/c. ábra). Unifaciális radiális levél jön létre akkor, ha a levél a színi oldal felé meghajlik, majd csőszerűen összezáródik, magába zárva az adaxiális epidermiszt. Ebben az esetben a szárhoz igen hasonló szöveti szerkezet figyelhető meg, kör alakban rendeződő szállítónyalábokkal. Unifaciális radiális levele a különböző hagymaféléknek (pl. Allium cepa [vöröshagyma]) van. A nősziromfélékre (Iris) a lemezes unifaciális levél a jellemző. Ezeknek a leveleknek az alsó része V-alakban összehajló (nyitott könyvhöz hasonló) szabályos bifaciális szerkezetet mutat, ami viszont a szártól távolabb eső szakaszokon összezárul (hasonlóan ahhoz, ahogy egy könyv becsukódik). Ekkor az érintkező színi epidermiszrétegek redukálódnak, eltűnnek és folytonossá válik a mezofillum. E levelek érdekessége, hogy bennük a szállítónyalábok kezdetben két sorban állnak (a két levélfélnek megfelelően), majd a levél csúcsa felé haladva a két oldal nyalábjai egymás közé csúsznak. Ekkor azonban a nyalábok kétféleképpen irányulnak: egyeseknek az egyik, másoknak a másik epidermisz felé néz a farészük. (Tehát csupán a metszeti kép alapján sem lehetne eldönteni, melyik a színi és melyik a fonáki oldal.) Az is előfordulhat, hogy a két levélfél nyalábjai nem egymás közé ékelődnek, hanem pont szembe kerülnek, majd összeolvadnak egymással, bikollaterális nyalábokat hozva létre.
A kétféle szövettani besorolás független egymástól. Tény, hogy a dorziventrális levelek az epidermisz szerkezete szempontjából többnyire bifaciálisak (pl. Nicotiana [dohány], Fagus [bükk], Nerium [leander], Nymphaea [tündérrózsa]) az ekvifaciális levél leggyakrabban izolaterális (pl. Galanthus [hóvirág], Viscum [fagyöngy], Sedum [varjúháj]) és az unifaciális levél szükségszerűen izolaterális is. Ennek ellenére előfordulnak pl. bifaciális, homogén mezofillumú levelek is (pl. Tradescantia [pletyka, mózeskosár], Stipa [árvalányhaj], Festuca [csenkesz]).
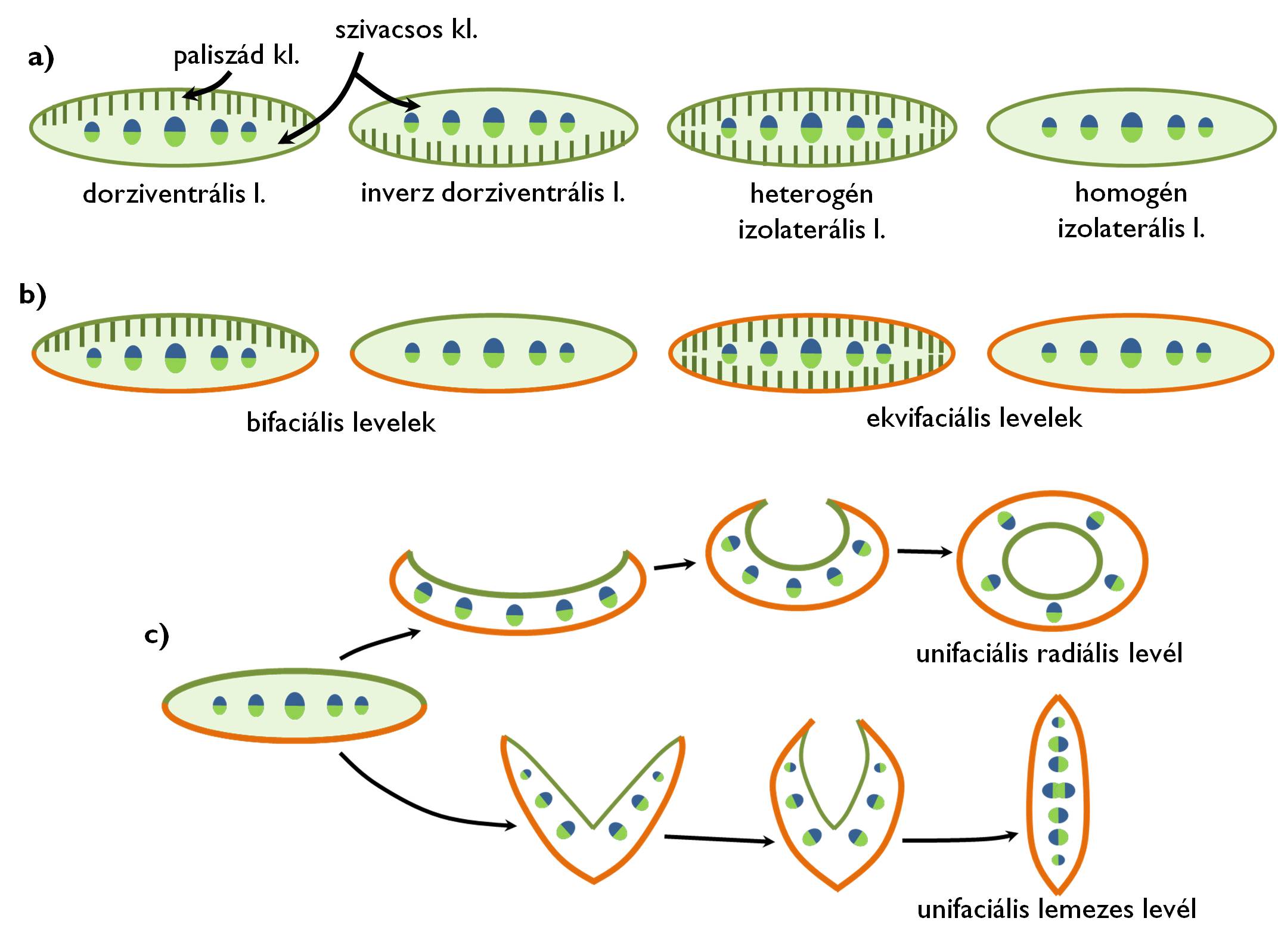
10. ábra. Levéltípusok a mezofillum (a) és az epidermisz (b,c) szerkezete alapján.
A hajtás ökológiai alkalmazkodásának lehetőségei
A növény hajtásának szöveti felépítése a különböző abiotikus környezeti tényezőktől függően megváltozhat. Ezeknek az evolúciós változásoknak két céljuk lehet: a növény által hasznosított tényezők maximális kihasználása illetve a szélsőséges környezeti paraméterek okozta stresszhatások kivédése, elkerülése.
A növényi anyagcsere (a fotoszintézis energiaigénye) szempontjából alapvető a fény szerepe. A növények jelentős része a maximális fénykihasználásra törekszik, amelynek egyik lehetősége, hogy lombleveleik eufotometrikusak, azaz a levéllemez mindig a fény beesési irányára merőlegesen fordul. A túlzottan napsütötte élőhelyeken ezzel szemben a sok fény mint stresszhatás jelentkezik. Az erős megvilágítás felmelegíti a leveleket, ami a párologtatás fokozódásához, majd kiszáradáshoz vezet. A fénystressznek azonban fiziológiai oldala is van: a fotoszintetizáló sejtekben nagy mennyiségben keletkeznek oxigéngyökök (reaktív oxigénformák), amelyek oxidálják, roncsolják a sejtalkotókat, és akár a sejtek pusztulását is okozhatják (oxidatív stressz). Éppen ezért az erősen napsütötte helyeken (pl. sziklagyepekben, homokpusztán) élő fajok igyekeznek kitérni a fénystressz elől, kisebb levélfelülettel, esetleg ún. pánfotometrikus levelekkel, amelyek élükkel fordulnak a nap felé, ezzel minimalizálva a lemezt érő sugárzás mennyiségét (pl. az Eucalyptus [eukaliptusz] fajok). (+info)
A nagy lombozattal rendelkező fás szárú növények esetében a lombkorona különböző pontjain különböző fényviszonyoknak kitett levelek találhatók: az erősen megvilágított felszínen fénylevelek, míg a lombozat belsejének fényszegényebb helyein árnyéklevelek. A fénylevelek lemeze többnyire nagyobb felületű, mezofilluma vastagabb, a fotoszintézis szempontjából aktívabb paliszád parenchima réteg aránya pedig jóval nagyobb (11. ábra). A kisebb felületű, vékonyabb (és nemritkán hosszabb levélnyelű) árnyéklevelekben ezzel szemben a szivacsos klorenchimaréteg a kiterjedtebb.
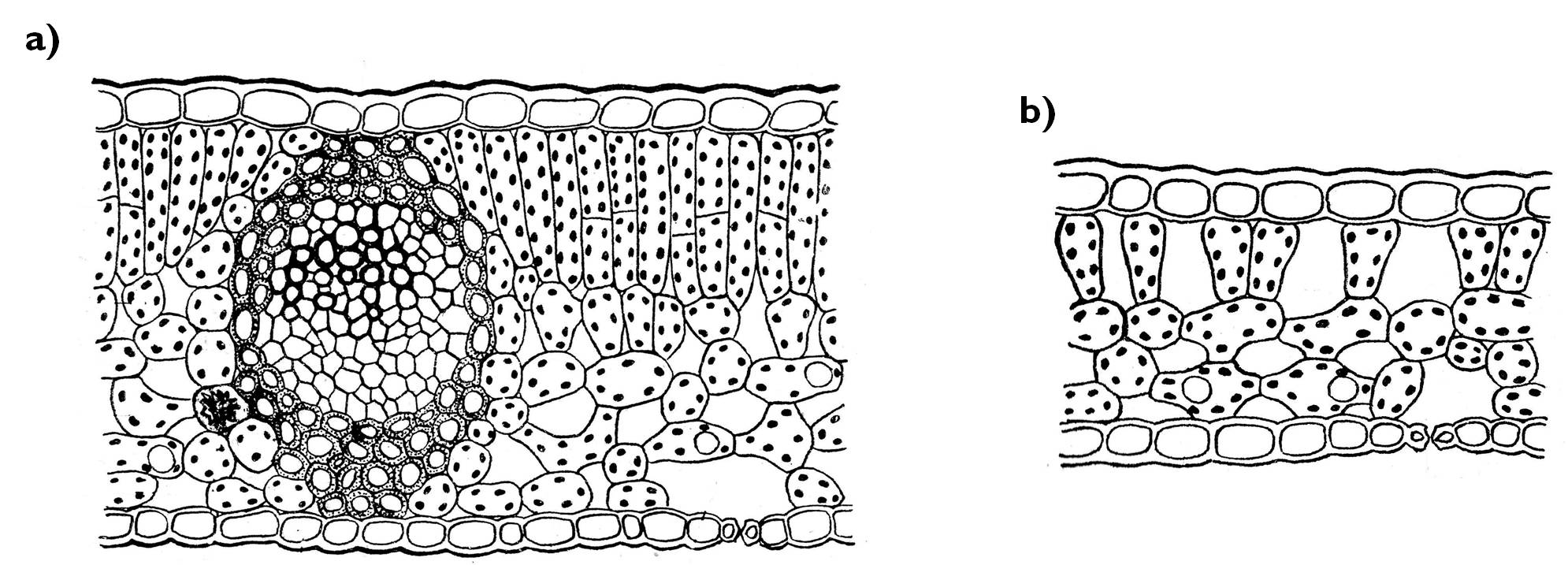
11. ábra. A fény- (a) és az árnyéklevelek (b) szerkezete.
(Sárkány és Szalai nyomán)
Különösen erős stresszhatást jelent – elsősorban a magashegységekben – a növényt érő nagy mennyiségű UV-sugárzás. Ellene a növények rövidebb internódiumok kialakításával, letörpüléssel védekeznek: ezeken az élőhelyeken gyakoriak a párnás növekedésű chamefiton fajok. Esetükben az egymást árnyékoló levelek segítik az erős sugárzás kivédését. A hajtást gyakran sűrűn beborítják az epidermális szőrök, amelyeket elhalt, levegővel telt sejtek alkotnak. A világos, fehéres szín a beeső fény nagy részét visszaveri, így az nem károsítja a hajtás belsőbb szöveteit.
A legfontosabb környezeti tényező, amelyhez a növények a hajtás morfológiájának és anatómiájának megváltoztatásával alkalmazkodnak, az a felvehető vízmennyiség. Habár a vízfelvétel folyamatát alapvetően a gyökér befolyásolja, a szervezet víztartalmát a növények elsősorban a levél (és a szár) vízleadása, valamint vízraktározás révén szabályozzák. Az „átlagos” vízellátottságú területeken élő növények (mezofitonok) leveleit (mezomorf levél) alapul véve, ezekhez képest vizsgálhatjuk a nedvesebb illetve szárazabb élőhelyek fajainak anatómiai bélyegeit (12. ábra). Az üde élőhelyeken (zárt lombkoronájú erdők aljnövényzetében, mocsár- és lápréteken) élő növényeket higrofitonoknak nevezzük (leveleik a higromorf levelek). E párás élőhelyeken élő fajok levelei vékonyak, kevéssé fejlett mezofillumúak (12/a ábra). A paliszád és a szivacsos klorenchima nehezen választható el egymástól, többnyire az egész levelet kiterjedt, tág üregű intercelluláris járatrendszer hálózza be. Mivel a mindig felvehető nedvességnek köszönhetően a sejtek turgora többé-kevésbé állandó, ez elegendő a növényi test megtartásához. Ennek köszönhetően e fajok hajtásaiban kevés a szilárdító szövet (rost, szklereida), így azonban igen hervadékonyak (pl. Impatiens [nebáncsvirág], Corydalis [keltike]). A levelek amfisztomatikusak, azaz sztómáik a levél mindkét felszínén előfordulnak, a zárósejtek pedig kiemelkednek a bőrszövet síkjából. A páratelthez közeli vízgőztartalmú élőhelyeken (pl. trópusi esőerdők aljnövényzetében) élő növények esetében könnyen gátolttá válhat a párologtatás folyamata. Ennek ellenére a gyökér ozmotikus szívóhatásából eredő gyökérnyomás végigpréseli a növény szervezetén a vizet és az ásványi anyagokat, ami cseppkiválásként jelenik meg a hajtás (elsősorban a levelek) felületén. A guttációnak nevezett jelenség a bőrszövet módosult sztómáin, a hidatódákon (lásd a 2. fejezetben!) keresztül zajlik. Ezek a szöveti elemek szintén egyes higromorf levelek sajátosságai (pl. Zantedeschia [kála]). A folyadékkal sűrűn borított levelek további jellemzője, hogy csúcsuk hosszan kihegyesedik (csepegtető csúcs), ami a folyadék elvezetését biztosítja a felületről. Ez a morfológiai jellemző megakadályozza a levél rothadását illetve egyes kórokozók megtelepedését.
A vízi környezet az itt élő hidrofitonok (vízi növények) speciális alkalmazkodását igényli. A hidromorf levél alapvetően kétféle lehet, a víztestbe merülő szubmerz levél (pl. Potamogeton [békaszőlő], Myriophyllum [süllőhínár]) vagy a vízfelszínen úszónatansz levél (Nymphaea [tündérrózsa], Nuphar [vízitök], Hydrocharis [békatutaj]). (+info) Mindkét típus sajátossága, hogy az egész felületen keresztül képes közvetlenül felvenni a vizet és ásványi anyagokat a környezetéből, ezért igen kisméretűek bennük a szállítónyalábok. A nyaláboknak különösen a xilem része fejletlen. A szubmerz levél a tápanyagok mellett a légzési ill. fotoszintetikus gázokat is diffúzióval veszi fel a környezetéből (sztómákat nem találunk bőrszövetében). Ennek következtében ezek a levelek igen nagy felületűek, erősen tagoltak, ám nagyon vékonyak, mezofillumuk redukált, sok esetben hiányzik (12/b ábra). A klorenchima szerepét ebben az esetben az epidermisz veszi át, ami miatt – kivételes módon – e fajok bőrszöveti sejtjei kloroplasztiszokat tartalmaznak. Az epidermiszen természetesen kutikulát sem találunk. Mivel a víz alatti hajtásrészek merevítését a víz felhajtó ereje végzi, minimális bennük a szilárdító alapszövet mennyisége. A natansz leveleket ezzel szemben a levélereket kísérő számos szklerenchima-rost és hipodermális helyzetű kollenchima-réteg tartja gerendázatszerűen feszesen (12/c ábra). E levelek mezofilluma kifejezetten vastag, egyrészt a több rétegű, oszlopos klorenchimaréteg miatt, másfelől pedig azért, mert a szivacsos parenchima intercellulárisai nagyméretűvé tágulnak, a szövet aerenchimává alakul. Ez a levegővel telt szövetréteg tartja a levelet a vízfelszínen. A tág üregekkel átjárt szövet szilárdságát a benne elszórtan elhelyezkedő szklereidák biztosítják. Sztómákat – magától értetődően – csak a színi oldalon találunk (episztomatikus levél). A levélnek ezt a felszínét vastag kutikula is borítja, ami vízlepergető tulajdonsága révén megakadályozza, hogy a levél átázzon és elmerüljön.
Abban az esetben, ha alacsony a talaj felvehető nedvességtartalma, a növény kétféle stratégiát folytathat: vagy párologtatását minimalizálja, vagy a lehető legtöbb vizet igyekszik felhalmozni szervezetében. A vízleadás csökkentésének lehetőségei közé tartozik a párologtató szervek, a levelek összfelületének csökkentése. A levelek redukálódhatnak pikkelyszerűvé (pl. Calluna [csarab], Tamarix [tamariska] fajok esetében) vagy levéltövisekké (ami többnyire levélszerű szár, fillokládium vagy kladódium kialakulásával jár együtt). Egyes nemzetségekben (pl. Euphorbia [kutyatej]) a száraz időszak beköszöntével a levelek elszáradnak, és a következő csapadék megérkeztéig a szár látja el a fotoszintézis feladatát. (+info) A levél párologtatásának csökkentését szolgáló szövettani szintű változások xeromorf levelek kialakulásához vezetnek (12/d-f ábra). Ezek a levelek többnyire kis felületűek, a hervadás okozta összeesést megakadályozandó sok szilárdító alapszövetet tartalmaznak. Bőrszövetüket – a bőrszöveti sejteken keresztül történő vízvesztés megakadályozása céljából – kutikularéteg borítja. A sztómák körül páratelt mikroklímát biztosító anatómiai változások figyelhetők meg. Ennek egyik lehetősége a szőrözöttség (ekkor a szőrök között megragadó vízgőz csökkenti a vízleadást). Gyakran elhalt, levegőt tartalmazó, világos színű szőrök borítják a felszínt, ami csökkenti a felület fényelnyelő képességét (azaz a párologtatást fokozó felmelegedés mértékét). A gázcserenyílások a bőrszövet többi sejtjének síkja alá süllyedve helyezkednek el (12/e ábra). Egyes fajok (pl. Nerium oleander [leander]) esetében a bőrszövet helyenként betüremkedik a mezofillumba, sztómakriptát hozva létre (12/d ábra). A gázcserenyílások ebbe a növényi szőrökkel bélelt, párával gyorsan telítődő üregbe nyílnak. A pázsitfűfélék (Poaceae) számos fajának jellegzetes bőrszöveti képletei a bulliform (ízületi) sejtek (12/f ábra), amelyek jellegzetes sejtfalvastagodásuknak köszönhetően vízvesztés esetén úgy változtatják alakjukat, hogy a levelek összepöndörödjenek (részletesen lásd a 2. fejezetben!). Ekkor a levél-cső belsejében alakul ki a magasabb vízgőztartalmú tér, ami csökkenti az ide nyíló sztómákon keresztül történő párologtatást.
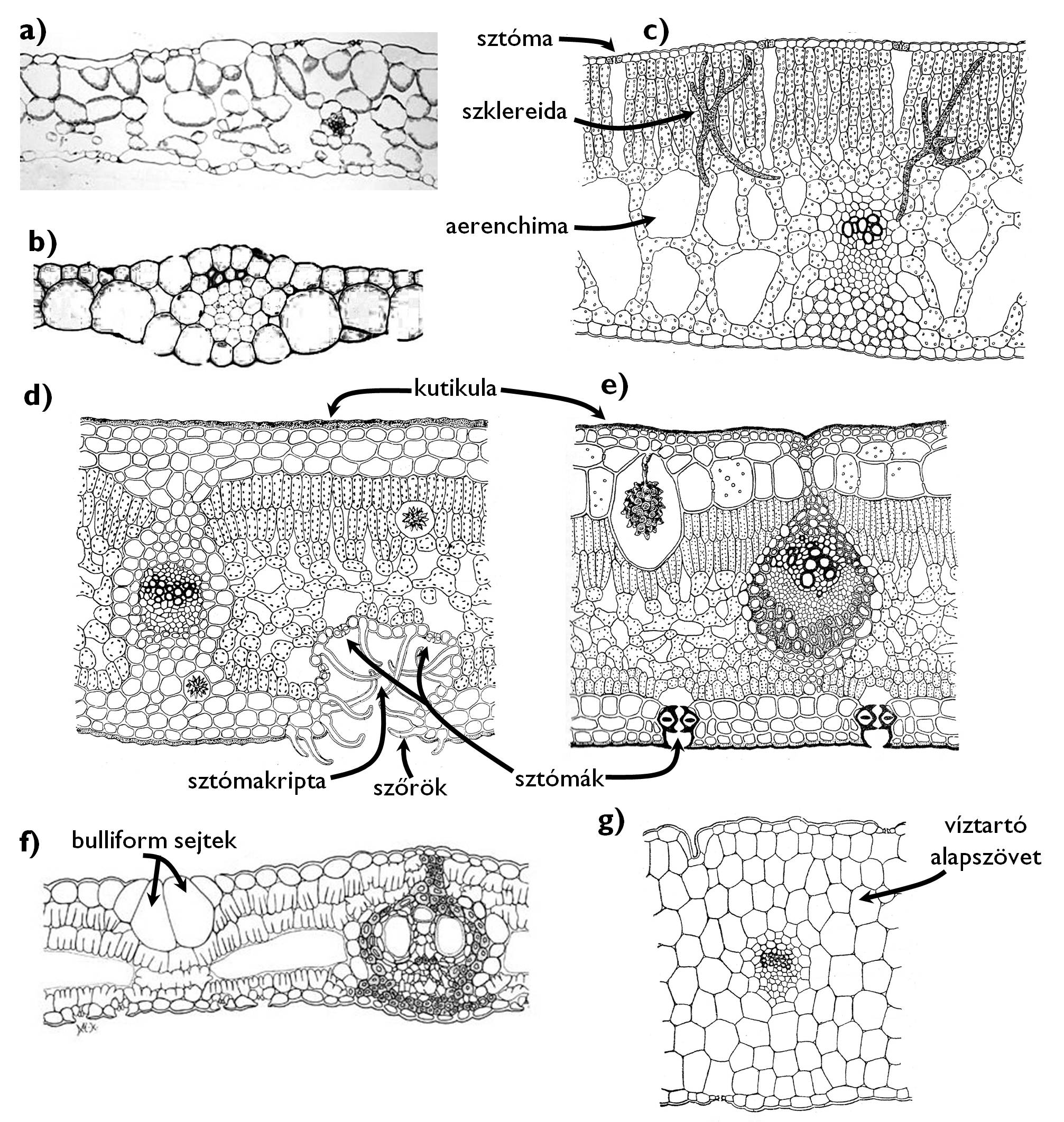
12. ábra. Különböző nedvességtartalmú környezethez alkalmazkodott levelek anatómiája.
a) higromorf levél (Impatiens); b) hidromorf-szubmerz levél (Elodea); c) hidromorf-natansz levél (Nuphar); d-f) xeromorf levelek (d: Nerium, e: Ficus, f: Phyllostachis); g) szukkulens levél (Sedum).
(a, b és f: internetes források nyomán: a, http://www.cas.miamioh.edu, b: http://esu.edu, f: http://www.eeob.iastate.edu/research/bamboo/characters/anatomy.html; c-e, g: Sárkány és Szalai nyomán)
A vízraktározás megvalósítása érdekében a xerofitonok szukkulens (pozsgás) szerveket hoznak létre. A víztartó alapszövet elhelyezkedhet gyökérben (Chlorophytum [csokrosinda]), hajtástengelyben (Euphorbiaceae [kutyatejfélék], Cactaceae [kaktuszok] család tagjai vagy a Adenium [sivatagi rózsa] nemzetség esetében), levéllemezben (Sedum [varjúháj], Sempervivum [kövirózsa]), levélnyélben (Zamioculcas [dűnefű], Rheum [rebarbara]). A szukkulens levelek jellegzetessége természetesen a nagyméretű, nyálkatartó vakuólumok tartalmazó víztartó alapszöveti sejtek jelenléte (12/g ábra). Természetesen a szukkulens levél esetében is megfigyelhetők a párologtatás csökkentésével együtt járó bélyegek: bőrszövetük viaszos, sztómáik besüllyedtek. A vízraktározás maximalizálása és a vízleadás csökkentése miatt e levelek relatív felülete (azaz a levelek felszín/térfogat aránya) kicsi, azaz ezek a levelek gömbszerű vagy hengeres megjelenésűek. (+info)
Nem a szárazság az egyetlen olyan abiotikus stressztényező, ami a xerofitonokra jellemző bélyegek kialakulásához vezet. Minden olyan környezeti hatás, ami megnehezíti a vízfelvételt (vagy fokozza a növény vízleadását) a korábban bemutatott anatómiai jellemzők megjelenésével jár (a kialakuló élettani szárazság miatt). Az erős napsugárzás, a hőstressz egyaránt fokozzák a párologtatást, míg fagy esetén a talajoldat víztartalmának jéggé alakulása nehezíti a vízfelvételt. A sóstressz (pl. szikes területeken) a talajoldat ozmotikus koncentrációjának megemelkedése miatt akadályozza a gyökér vízfelvételét. Ennek megfelelően például a tajga öv vagy a tundrák növényei és a tengerparti zónákban illetve a kontinentális szikeseken élő fajok egyaránt a szárazsághoz történő alkalmazkodásra utaló morfológiával és anatómiával rendelkeznek.
A napos, meleg területeken élő növények vízmegőrzést szolgáló adaptív fiziológiai jellemzője, hogy nappal a párologtatás csökkentése céljából bezárják gázcsere-nyílásaikat. Ennek azonban az a következménye, hogy a fotoszintézis fényszakasza szempontjából fontos időszakban erősen lecsökken a levelek intercellulárisainak szén-dioxid-koncentrációja. Ennek a problémának a megoldására az evolúció során alternatív fotoszintetikus folyamatok jöttek létre. A növényvilágban általánosan elterjedt mód a C3-as fotoszintézis, amely nevét arról kapta, hogy a CO2 megkötésével egy három szénatomos vegyület (glicerinsav-3-foszfát) jön létre, ami egy többlépéses körfolyamatban (sötétszakasz, Calvin-ciklus) a fényszakaszban megtermelt ATP és NADPH segítségével glükózzá alakul (részletesebben l. a kloroplasztisszal foglalkozó fejezetben!). A C3-as növényekben a fény- és a sötétszakasz időben egyszerre (nappal), egyazon sejt kloroplasztiszának belsejében megy végbe. A meleg, száraz környezetben élő növények egy jelentős része ehelyett ún. C4-es fotoszintézist folytat, aminek az a lényege, hogy a CO2 megkötése és glükózzá alakítása térben elkülönülve, két külön sejttípusban megy végbe. A levél mezofillumsejtjeiben kerül sor az első lépésre, ahol a megkötött CO2 egy négy szénatomos vegyületté (almasavvá, oxálecetsavvá vagy szerinné) alakul. Innen ez a négy szénatomos vegyület átkerül a levélereket övező nyalábhüvelyparenchima-sejtekbe, ahol felszabadul belőle a CO2, és a „hagyományos” C3-as módon, a Calvin-ciklusban glükózzá alakul. A C4-es fotoszintézis előnye, hogy a nyalábhüvelyparenchima-sejtekben a CO2-koncentráció olyannyira megemelhető, hogy zárt sztómák mellett is hatékony szervesanyag-termelés válik lehetővé.
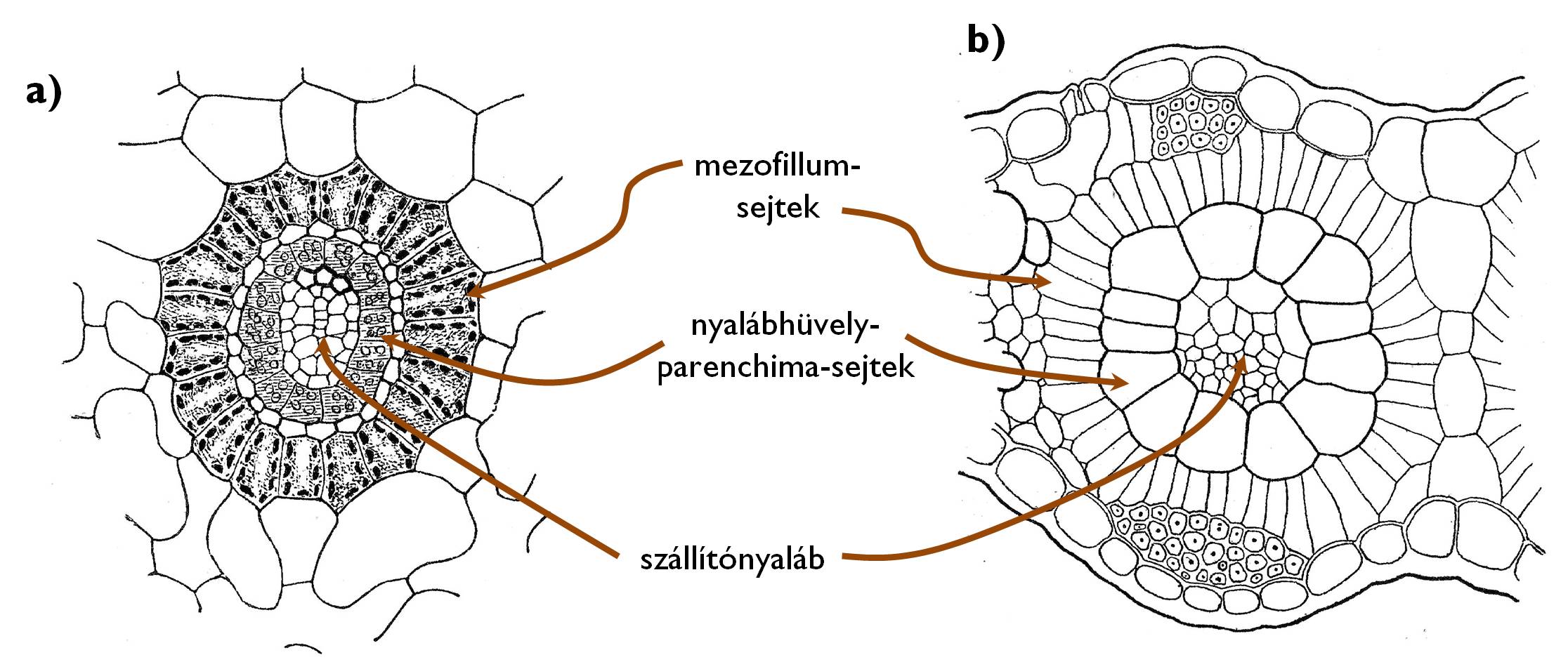
13. ábra. A Kranz-anatómia. (a: Cyperus papyrus [papirusznád], Panicum miliaceum [köles])
(Habelrandt nyomán)
A C4-es fotoszintézis mint élettani adaptáció anatómiai változásokkal is jár. A levelekben a levélér körül szabályos, koncentrikus gyűrűkként figyelhetők meg a nyalábhüvelyparenchima-sejtek, majd ezeken kívül hasonlóan rendezetten a mezofillumsejtek rétegei (Kranz-anatómia) (13. ábra). Emellett a C4-es fajok egy jelentős részében mikroanatómiai változások is bekövetkeznek. A mezofillumsejtek gránumos kloroplasztiszaiban nem találunk keményítőszemcséket, mivel nem megy bennük végbe glükózképzés. (A gránumok jelenléte arra utal, hogy a fényszakasz itt végbemegy, mivel a C4-es folyamat esetében a CO2 megkötése itt igényli a megtermelt ATP és NADPH nagy részét.) Ezzel szemben a nyalábhüvelyparenchima-sejtek színtesteiben nem találunk gránumokat (mivel bennük nem zajlik a fotoszintézis fényszakasza), viszont előfordulnak a megtermelt glükóz raktározására szolgáló (primer) keményítőszemcsék. Ezt a jelenséget nevezzük a C4 -es növények plasztiszdimorfizmusának.
Ellenőrző feladatok
Ide jó lenne a feladatokat feltölteni
Irodalomjegyzék
Beck CB (2010): An Introduction to Plant Structure and Development: Plant Anatomy for the Twenty-First Century. 2. kiad. Cambridge University Press, Cambridge,
Biebl R, Germ H (1967): Praktikum der Pflanzenanatomie. 2. kiad., Springer-Verlag, Wien-New York.
Braune W, Leman A, Taubert H (1967): Pflanzenanatomisches Praktikum. VEB Gustav Fischer Verlag, Jena.
Cutler DF, Botha T, Stevenson DW(2008): Plant Anatomy: An Applied Approach. Wiley-Blackwell, Oxford.
Erős-Honti Zs (2012): A kertészeti növények alaktana. Egyetemi jegyzet. Budapesti Corvinus Egyetem Kertészettudományi Kar, Bp.
Evert RF, Eichhorn SE (szerk.) (2006): Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body – Their Structure, Function, and Development, 3. kiad., Wiley-Interscience, New Jersey.
Gyurján I (1996): Növényszervezettan. Egyetemi jegyzet. ELTE Eötvös K., Bp.
Haberlandt G (1924): Physiologische Pflanzenanatomie. Verlag von Wilhelm Engelmann, Leipzig.
Jacob F, Jäger EI, Ohmann E (1985): Botanikai kompendium. Natura, Bp.
Kárpáti Z, Görgényi L, Terpó A (1968): Kertészeti növénytan I. Növényszervezettan. Mezőgazdasági Kiadó, Bp.
Mauseth JD (2008): Plant Anatomy. The Blackburn Press, New Jersey.
Sárkány S, Haraszty Á (1995): Növényszervezettan biológia-kémia, biológia ill. pszichológia-biológia szakos hallgatók részére. Egységes jegyzet. Nemzeti Tankvk., Bp.
Sárkány S, Szalai I (1966): Növénytani praktikum I. Növényszervezettani gyakorlatok. 3. kiad., Tankvk., Bp.
Wilson K, White DJB (1986): The Anatomy of Wood: its Diversity and Variability. Stobart & Son Ltd., London.
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028