A növényvilág diverzitása, növényfajok és populációk
Szerző: Höhn Mária
Biológiai diverzitás a Földön
Tudjuk mennyire változatos a földi élet, hányféle élőlény kötődik a különféle életterekhez. Eme változatosságra, sokféleségre használt fogalom a biológiai diverzitás.
A Föld története során az élővilág diverzitása fokozatosan alakult ki és korszakról korszakra sokat változott. Voltak geológiai korok, amelyek nagy fajkihalásokkal, mások fajok tömeges szétterjedésével jellemezhetők. A diverzitás változása nagymértékben függ földtörténeti tényezőktől, a geológiai korok klimatikus jellemzőitől, paleogeográfiai, geomorfológiai tényezőktől, az egyes élőlénycsoportok genetikai adottságától, alkalmazkodó képességétől.
Azt mondjuk, hogy a biológiai diverzitás (biodiverzitás) a geodiverzitással szorosan összefügg, hiszen minél változatosabb a környezet és minél többféle élőhely alakul ki, annál változatosabb lehet az azt benépesítő élővilág is. A geodiverzitás a környezeti tényezők sokféleségéből adódik.
Ha föltesszük azt a legegyszerűbb kérdést - Hány faj él a Földön?- nehéz egyértelmű választ adni. Először tisztáznunk kell azt, hogy ismerjük-e a Földön élő fajok mindegyikét?
A válasz egyértelműen nem. Bár a megismerés folyamata régen kezdődött és már az ókorban számos növény és állatfaj leírását közölték, mai napig csak becslések vannak a ma élő fajok számáról. Az 1.táblázat jól szemlélteti azt a tényt, hogy a megismert és becsült fajok száma között az egyes élőlénycsoportok összehasonlításában mekkora különbségek vannak.
1. Táblázat: Az ismert és becsült fajok száma a Földön (Cristea és Denaeyer, 2004)
-
Rendszertani
csoportokLeírt fajok
számaLehetséges fajok
száma a FöldönA leírt fajok
%-ban kifejezveVírusok
4000
400 000
1
Baktériumok
4000
1 000 000
0,4
Gombák
72 000
1 500 000
5
Egysejtűek
40 000
200 000
20
Algák
40 000
400 000
10
Mohák
14 000
30 000
47
Edényes növények
250 000
300 000
83
Gyűrűsférgek
25 000
400 000
6
Rákfélék
40 000
150 000
27
Pókszabásúak
75 000
750 000
10
Rovarok
1 000 000
8 000 000
13
Puhatestűek
100 000
200 000
50
Gerincesek
44 000
50 000
88
Más csoportok
115 000
250 000
46
Összesen
1 800 000
13 600 000
13
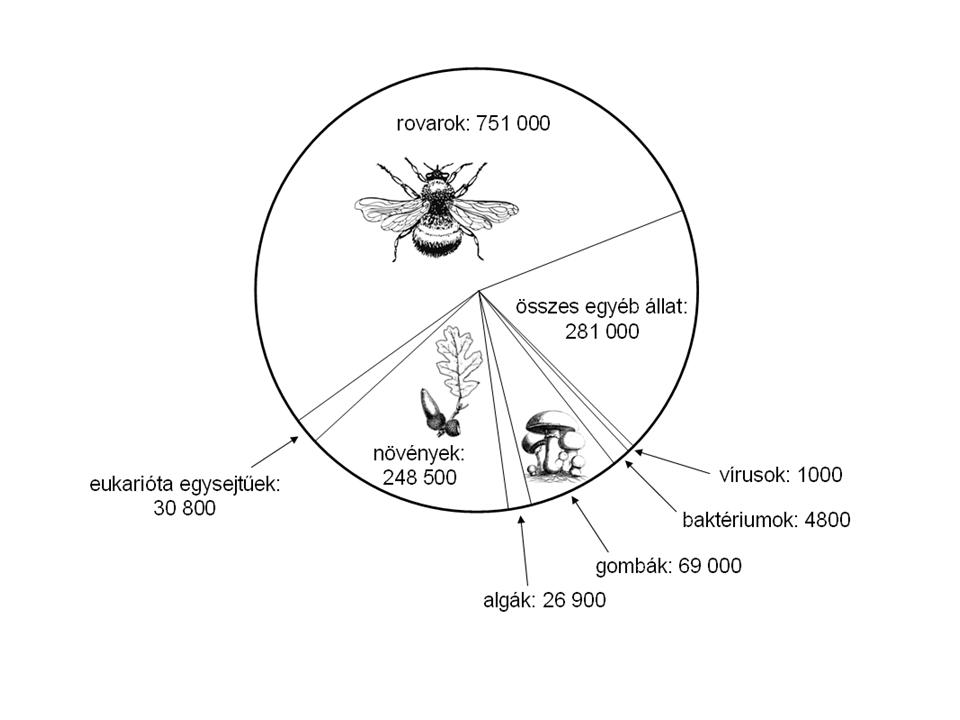
1. Ábra. A földi biodiverzitás eloszlása rendszertani kategóriák szerint (Standovár és Primack 2001 nyomán)
Legjobban a magasabbrendű növények és állatok csoportjait ismerjük, míg a rovarvilág becslések szerint 8 millió faja közül mindössze 13% ismert a tudós társadalom számára. Fenti szerzőkre hivatkozva azt mondhatjuk, hogyha több mint 13 millió faj a becsült érték, akkor a számunkra ismert 1,8-2 millió faj csak az összfajszám 13% -át teszi ki.
Az ismert fajok számának rendszertani csoportok szerinti eloszlását, arányait az 1. ábra mutatja.
Egyéb irodalmi adatok (Mora és mtsi. 2011) szerint körülbelül 8,7 millió faj élhet jelenleg a Földön (± 1,3 millió) és ebből a tengerek világában 2,2 millió faj található. (Részletesen l. a következő honlapon: http://www.diversitas-international.org/activities/research/biogenesis)
Akárhogyan is számoljuk, megállapítható, hogy a tízmilliós nagyságrendű fajdiverzitás mellett a magasabbrendű növények, melyek többségét vélhetőleg már ismerjük, mintegy 280-300 ezer faja a Földi diverzitás kis hányadát képviseli.
A magasabbrendű növények, nyitvatermők és zárvatermők, melyek maggal terjednek a szárazföld meghódításában voltak sikeresek. A mag mint nyugalomra képes ivaros szaporító szerv biztosította számukra azt a lehetőséget, hogy csírájuk a mag belsejében szárazföldi körülmények között akár a kedvezőtlen időszakban is életben maradjon és az új élet fejlődése, a csírázás csak a külső körülmények alkalmassá válása esetén indulhasson el (részletesen l. a 4. fejezetben!). Mind a terjedésben, mind a csírázás első kezdeti szakaszában a mag funkcionális struktúrái segítik az embriót.
Természetesen, a fajok, így a növényfajok léte átmeneti jelenség, különböző korokban más-más fajok éltek és élnek, így a diverzitás a földi élet során mindig is változott. Az evolúció gondolatának bevezetésével, Darwin óta tudjuk, hogy a diverzitás változását az evolúció motorja is meghatározza.
Borhidi Attila (2009) hazai és külföldi kutatási eredményekre alapozva, a növényvilág evolúciójában négy fontos stádiumot különít el. Ezek valójában olyan fejlődési szakaszok, amelyek kezdeti időszakát az új típusok, új fajok viszonylag gyors és gazdag megjelenése jellemzett:
- Az őstenger élővilágának kialakulása: (földtörténeti ókor – kambrium kora). Szinte minden készen áll a teljes földi élet evolúciójávoz (9 algatörzs és a gerincesek kivételével 17 állattörzs)!
- A növények partraszállása, (devon kor, a földi légkör oxigéntartalma 10%-ra növekszik). Kialakul az ózonpajzs, megszűnik az erős UV sugárzás a földfelszínen. A szállítószövetek kialakulásával megjelenik a vízszállítás képessége → hajtásos növények (valódi szövetes felépítés).
- A nyitvatermő növények szétterjedése, (devon vége, karbon eleje), a mag megjelenése.
- A zárvatermők robbanásszerű megjelenése, (perm végétől: virágba borul a szárazföld)
A termés és megporzás sokfélesége, termés terjesztés. Közben elterjednek az állatok is hiszen:
„Az állatvilág − az embert is beleértve − a növényvilág vendégeként éli földi életét. „
A diverzitás jellemzésére használhatók a diverzitási mutatók, melyek számszerűsítve fejezik ki valamely terület vagy élőhely élővilágának sokféleségét.
Legtöbbet használt mutatók az α, β és γ-diverzitás
α – egy élőhely fajainak száma (taxonszám)
β – élőhely sorozat taxondiverzitása, viszonyszám, hasonlósági mutató
γ – egy nagyobb régió, pl. egy egész ökoszisztéma diverzitása, egy táj, egy biom sokfélesége
A béta diverzitás egyazon nagyobb régió több területének –egy transzekjének- összehasonlítását teszi lehetővé. Legegyszerűbben:
β1 – γ/ α1
β2 – γ/ α2
A diverzitás jellemezhető a közösséget alkotó fajok illetve egyedeik relatív gyakoriságából származtatott mutatókkal is:
Ilyen egyszerűbb diverzitás függvény az ún. Shannon-Wiener index (H)
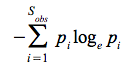
- ez az információelméletből „kölcsönvett”, egyik leggyakrabban használt index
ahol: pi – i-ik faj relatív gyakorisága a közösségben
vagy a Simpson index
mekkora a valószínűsége annak, hogy egy felbukkanó egyed ugyanahhoz a fajhoz tartozzon, mint egy korábban felbukkanó egyed.
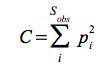
ahol 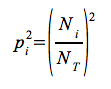 és végül maga az index:
és végül maga az index: 
Ni – i-ik fajhoz tartozó egyedek száma
NT – a mintába tartozó összes egyed száma
A diverzitás összehasonlítását (pl. két élőhely diverzitását) teszik lehetővé olyan indexek, mint a Jaccard vagy a Soerensen mutatók. (szövegdoboz !)
Fontos megjegyezni, hogy a fajdiverzitáson alapuló mutatók a fajszám mellett az egyes fajok előfordulási gyakoriságát, a fajok eloszlását, és egyenletességét is figyelembe veszik. Így például két élőhely összehasonlításában diverzebb élőhelynek számít az, ahol az egyenetlenség a legnagyobb, vagyis a fajok megjelenési esélye hasonló vagy közelítő értékű.
A diverzitásnak a taxondiverzitáson túl, más fontos szintjei vannak, mint az
-
egyedi szintű:
- morfológiai diverzitás
- genetikai diverzitás: allélgyakoriságokból számított diverzitás (α diverzitás)
-
faji szintű:
- egyedek és a populációk sokfélesége (l. korábban)
-
ökoszisztéma szintű:
- fajszám / terület: számszerűsíthető különböző mutatók mentén
- filogenetikai sokféleség, a rokoni kapcsolatok különbözősége
- ökológiai funkciók sokfélesége (ökológiai diverzitás)
Tágabb értelmezésben pedig beszélünk:
- életformák
- élőhely típusok számának
- alapkőzet és talaj, domborzat, vízhálózat
- tájképi elemek
diverzitásáról.
Miért is fontos a diverzitás? Miért jó, ha magas egy terület biológiai és geodiverzitása egyaránt?
Mert:
- több, változatosabb élőhely több élőlényt tud eltartani;
- a nagyobb élőhely diverzitás nagyobb biodiverzitást hoz létre;
- ha több élőlény van, akkor közöttük többféle, sokrétű kapcsolat alakul ki;
- ha többféle kapcsolat alakul ki és azok többnyire állandósulnak, akkor rendszerszerűen működnek;
- a rendszer stabilabbá válik, mert minél bonyolultabb és minél több komponensből áll, annál nehezebben ingatható meg;
- kialakulnak azok a feltételek, amelyek a rendszer önfenntartó képességét biztosítják!
Következésképpen:
A nagyobb diverzitás nagyobb stabilitást eredményez, és ez a természet működési rendje!
A diverzitás az evolúció folyamán formálódott, és igazolódott az a tény, hogy az ökoszisztémák minél nagyobb diverzitásúak, fenntartásuk annál kevesebb energiát igényel. A diverzitás megőrzése egy sikeres túlélési stratégiát jelent a legtöbb ökoszisztéma számára.
Természetesen a diverzitás növekedése is véges, a nagy fajszámú ökoszisztémák belső kapcsolatrendszere önszabályozóan korlátozhatja a diverzitás káoszba vezető növekedését (2. Ábra).
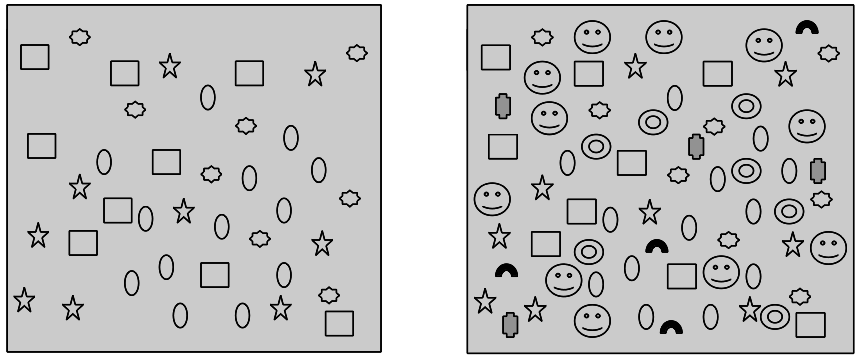
2. Ábra. Egy élőhely különböző fafajokhoz tartozó egyedeinek (ikonok) eloszlása és sűrűsége (Standovár T. nyomán) esetén előfordul, hogy a fajok egyedei a nagy egyedsűrűség miatt egymástól távol kerülnek és reprodukciós akadályok lépnek föl. Ekkor az egyedszám gyarapodás megáll és érvényesül az ökoszisztéma önszabályozó mechanizmusa.
Mivel az emberiség kialakította ökoszisztémák működése is a természetes közösségekéhez hasonló, a termesztési gyakorlatban is mindinkább hangsúlyossá válik a diverzitási szempontok figyelembevétele. A mai mesterséges öko-rendszerekben az élőanyag mennyisége (biomassza) és minősége (biodiverzitás) nagyságrendekkel elmarad a régi természetes öko-rendszerekétől (Vida 1996). Az intenzív agrárrendszerek világméretű terjedése azonban instabilitáshoz vezetett. A fajtaszám-csökkenés, illetve néhány kultúrfaj és fajta egyeduralkodóvá válása az egész világgazdaságot sebezhetővé tette, és a nemesítési alapanyagok eltűnésével az újabb, alkalmasabb fajták előállításának lehetősége is veszélybe került. Azt mondhatjuk, hogy az agro-biodiverzitás megőrzése napjainkban szükségszerűség és elengedhetetlen az emberi jólét és biztonság megőrzéséhez és fenntartásához.
Migráció és diszperzió jelensége a növényvilágban
A természetes fajok élettartama változó. A Föld története során fajok tűntek el, pusztultak ki mások megjelentek. A kipusztulás egy természetes folyamat. Az emberi hatás következtében azonban ma már a fajpusztulás mértéke rendellenesen felgyorsult és tudjuk, hogy messze meghaladja a természetes értéket. A fajok kihalásának okai többfélék, a túlhasználattól az élőhelyek megszűnéséig sok tényező okozhatja a fajok idő előtti kipusztulását.
Történetük során a fajok vándorolnak, ami a fajok areájának (elterjedési területének) megváltozásával jár. Ezek a változások hosszú idő alatt zajlanak és ennek eredményeképpen egy faj areája tágulhat (nőhet), szűkülhet, vagy el is tolódhat egy földrajzi grádiens mentén.
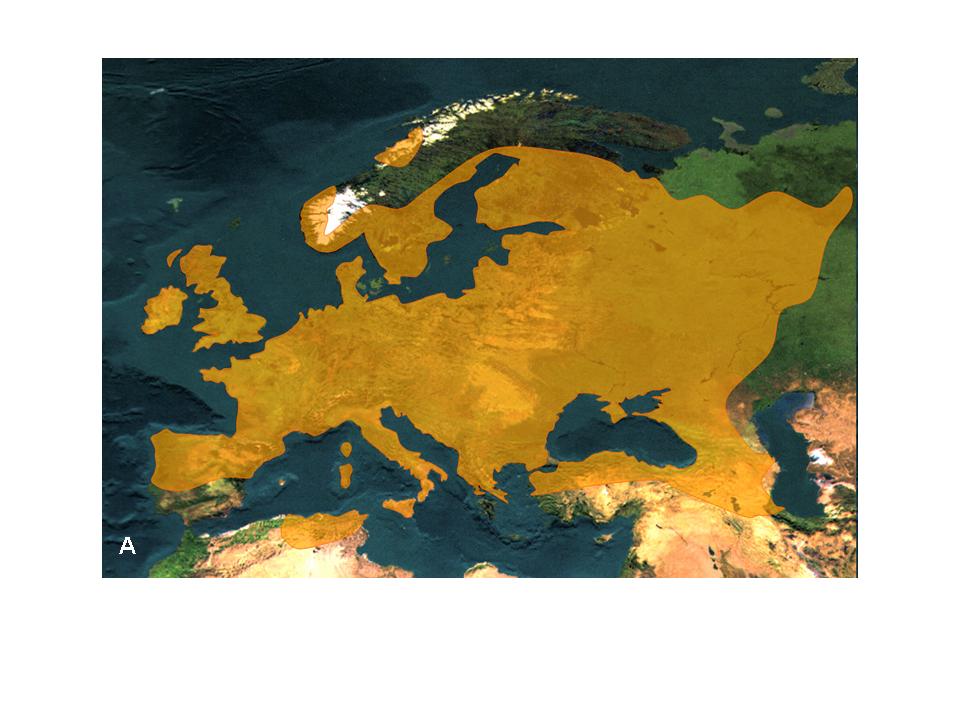
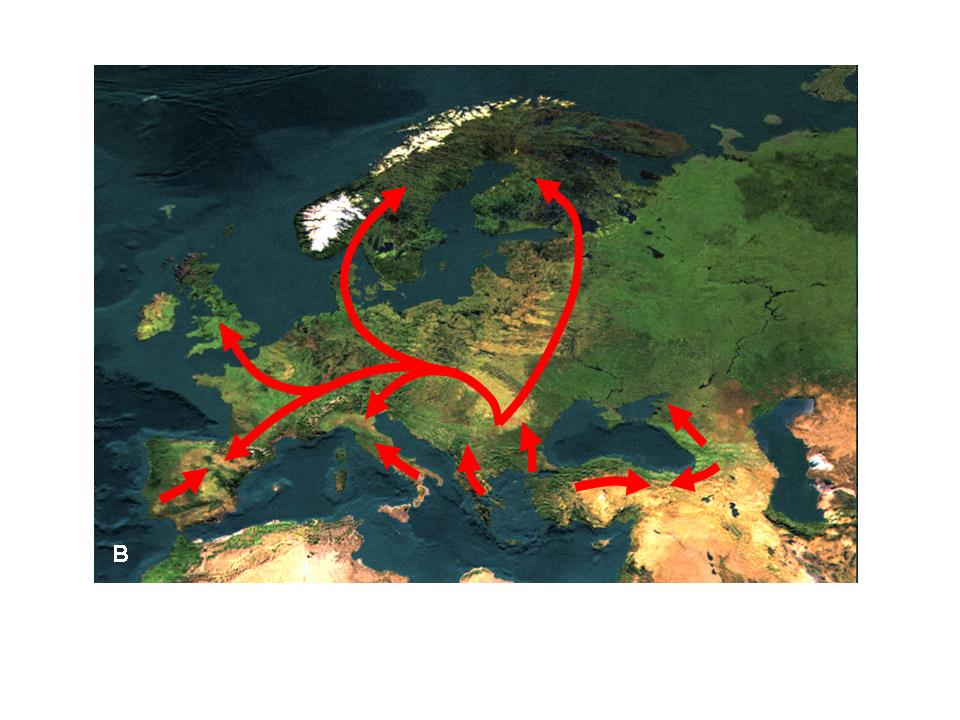
3. Ábra. Az enyves éger (Alnus glutinosa) areatérképe (A) és poszglaciális migrációjának fontosabb útvonalai (B) (Hewitt 2001 nyomán)
Miért vándorolnak a fajok? Migrációs kényszer hatására, amely a kipusztulás fenyegetettsége miatt áll fenn. A megváltozó körülmények hatására fajok areájuk elhagyására kényszerülhetnek (pl. jégkorszak beköszönte) miközben vagy eltűnnek vagy új területeket kolonizálnak (3. Ábra). Ha egy faj elterjedési területe teljesen megváltozik, és areája új földrajzi térségek mentén körvonalazódik, akkor beszélünk areán túli migrációról. Általában azonban a faj areáján belül is „mozoghat”, így areája feldarabolódhat és/vagy átrajzolódhat. Az erdeifenyő (Pinus sylvestris) a jégkorszak elmúlta után északabbra húzódott, és a jég által felszabadított területeket hódította meg, ahol ma is a tajgaerdők egyik legfontosabb erdőalkotó fafaja (4. Ábra). Eközben Európa középső részén areája beszűkült, egykori területeit a lombos fajok, a hegyvidéken pedig a lucosok foglalták el. Az erdeifenyő kipusztulása Dél- és Közép-Európában a felmelegedést követően azért nem következett be, mert tág tűrőképessége folytán populációi szélsőséges élőhelyeken, lápokban, sziklás meredek oldalakon fenn tudtak maradni (5. Ábra).
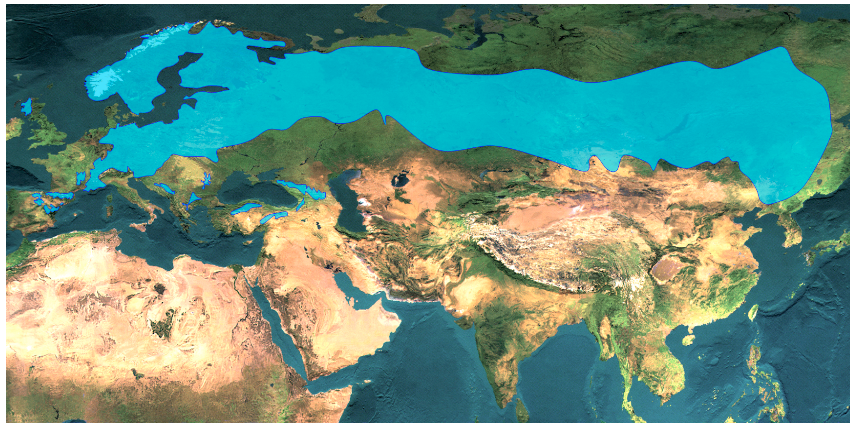
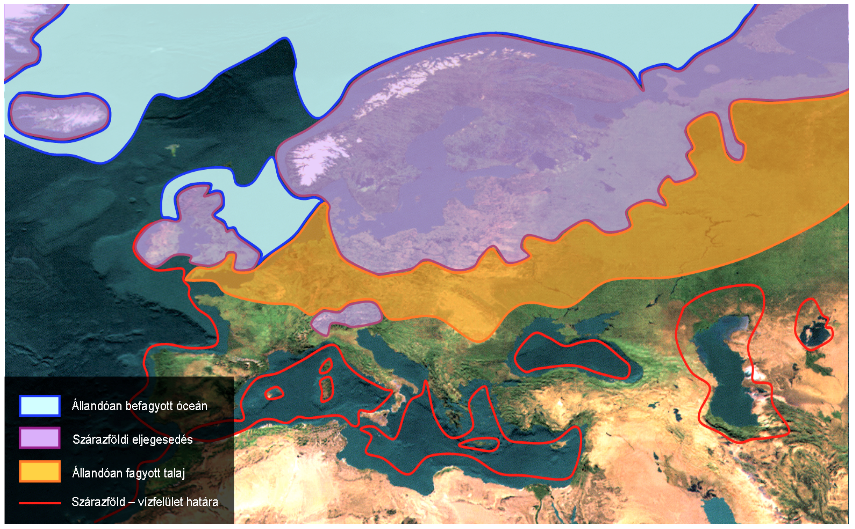
4. Ábra. Az erdeifenyő (Pinus sylvestris) jelenlegi areatérképe (Euforgen nyomán) (A). A szárazföldi jégtakaró kiterjedése és az állandóan fagyott talaj határa az utolsó eljegesedés hidegmaximuma alatt (B) (Láng, Bedő és Csete 2003). A fenti két térkép alapján látható, hogy az erdeifenyő mai elterjedési területének nagy részét a jégkorszaki maximum idején kiterjedt jégpáncél borította.



5. ábra. Az erdeifenyő (Pinus sylvestris) szélsőséges termőhelyei Dél- és Kelet Európában. A: meszes homokon Fenyőfőn (Bakonyalja); B: a tusnádi Mohos tőzeglápban (Csomád hegység); C: a ’Poiana Stampei’ tőzeglápban (Kelemen havasok)
Kisléptékű „elmozdulás” is jelenthet sikeres túlélést. Például a jégkorszak elmúltával számos faj Európában, így a középhegységekből is a közeli magasabb hegyvidékre kényszerült, mert helyüket melegkedvelő fajok foglalták el. A hegyekről olvadó hósapka zsugorodásával azonban, új csupasz felszínek jöttek létre, és ez új életteret biztosított számukra, mintegy menedéket nyújtott ezeknek fajoknak.
Van olyan alapkőzet is, mint a dolomit, amely fizikai-kémiai tulajdonságaival olyan változatos felszíneket volt képes létrehozni, amely mikro-élőhelyek kialakulását eredményezte. Így a hidegebb korok fajainak egyes egyedei ezeken a mikro-menedékekben maradhattak fenn. Ezzel magyarázzuk például azt, hogy északi dolomit lejtőinken, ma is élnek olyan növények, melyek legközelebb már csak az Alpok-Kárpátok magashegységein találhatók meg, pl. Phyteuma orbiculare (gombos varjúköröm), Primula auricula (medvefül kankalin) (6. Ábra).

6. Ábra. Phyteuma orbiculare - gombos varjúköröm (A) és Primula auricula – medvefül kankalin (B)
A Dunántúli Középhegység dolomitfelszíneinek maradványflórája igen fajgazdag. A jégkorszak hosszú ideje alatt rövidebb-hosszabb felmelegedések voltak. Az életben maradt populációk elszigetelődve a korábbi elterjedési területükön élő egyedektől hosszú idő alatt új fajokká, alfajokká alakulhattak át. Ilyen például a pilisi len (Linum dolomiticum), mely a Pilis hegységben (mai egyetlen élőhelye a Kis-Szénás) az északi fekvésű dolomitlejtőkön maradt fenn (7. Ábra.). Ma már Európa egyik érdekes és értékes bennszülött fajának tekintjük, melynek története a pleisztocéni időkre vezethető vissza. Legközelebbi rokonának a Linum elegans-t tekintik, mely jóval délebben a görögországi mészkőhegyek napos szikláin él.

7. ábra. Linum dolomiticum – pilisi len (Udvardy László felvétele)
Populációk
Egy faj egyedei csak akkor képesek termékeny utódok létrehozására, ha a különnemű egyedek találkoznak egymással, vagyis egy élőhelyen együtt vannak, és akadálytalanul ivarosan szaporodnak. Az egy fajhoz tartozó egyedek, melyek egy élőhelyet közösen népesítenek be, populációkat építenek fel. Így a fajt az élőhelyen (habitat) a populáció képviseli. A faj areája mentén rendszerint számos populáció él, és ezek a számukra alkalmas élőhelyeket benépesítik be.
Tehát: Az areán a fajt az élő populációk összessége alkotja.
Populáció fogalma
Egy faj egyedeinek azon csoportja, amelyek térben és időben együtt fordulnak elő, ezért egyedei között folyamatos génkicserélődés, génáramlás zajlik. A populáció egy valós szaporodási közösség. Egy adott élőhelyen egy fajt rendszerint egy populáció képvisel. Nem egyszerűen az egyedek sokaságáról van szó, hanem egy olyan egyedcsoportról, amely meghatározott struktúrával és funkcióval írható le.
Diszperzió (szóródás)
Tudjuk, hogy a helyváltoztatásra képtelen növények vándorlása „elmozdulása”, nem aktív helyváltoztatást jelent. Valójában arról van szó, hogy a nagy területeken elszóródó magvaik közül azok, amelyek éppen „jó helyre” kerülnek és csíráznak, biztosítják a populáció túlélését és az új élőhelyek benépesítését.
Ha egy faj vándorol (migráció), megváltozik az areája. A populáció viszont szóródik (diszperzió) a terjesztőképletei által. A terjesztőképletek vagy propagulumok (magok, spórák, vegetatív képletek), elszóródva új élettereket népesíthetnek be, ezzel biztosítva a faj fennmaradását és új élőhelyek meghódítását. A diszperzió tehát egy populációszintű esemény, és a populációdinamikai folyamatok eredményeképpen a növényfajok vándorlása zajlik.
A növényi populációk jellemzői: általános és specifikus bélyegek
Ha bármely növény- vagy állatfaj populációját vizsgáljuk néhány általános jellemzőjével írhatjuk le.
Ilyenek:
a) az egyedszám: hány egyed alkotja a populációt,
b) a születési ráta: az egységnyi idő alatt született utódok száma
c) a mortalitási ráta: az egységnyi idő alatt elpusztult egyedek száma
Kétféle értéke lehet: a biológiai mortalitás, amely a populáció specifikus élettartamából adódik és az ökológiai mortalitás. Utóbbi egy olyan érték, amely erősen függ a környezettől. Ha fokozódik a populációra ható környezeti stressz hatása (kevesebb táplálék, víz, sok versengő partner vagy éppen ellenséges populáció), az ökológiai mortalitási érték növekszik.
d) koreloszlás: az egyes generációk aránya (8. Ábra)
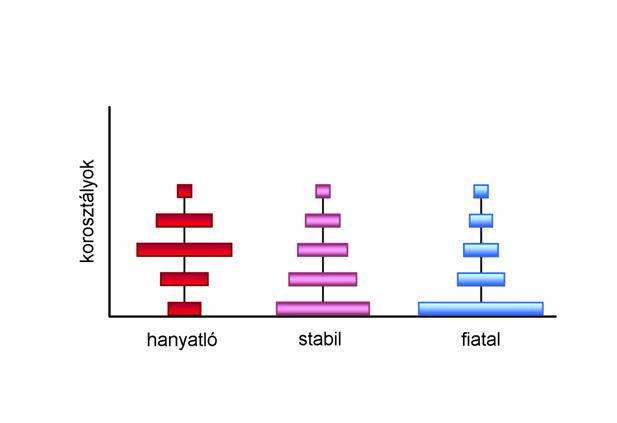
8. ábra. Öregedő (hanyatló), stabil és fiatal populációk modelljei.
e) növekedés: Nt+1=Nt + B - D +I - E
Nt – 0 órában mért, kiinduló egyedszám; B- született egyedek (born); D – elpusztult egyedek(dead); I – bevándorlók (imigrants); E – elvándorlók (emigrants)
f) reguláció (szabályozás)
A populációk saját túlélésük érdekében az egyedszám szabályozására kényszerülnek.
Populáción belül a szaporodási képesség közvetlen következményeként az utódok létrehozása többé-kevésbé folyamatos és ez az esemény leírható az „r”értékkel, melyet reprodukciós rátának nevezünk.
dN/dt = rN , ahol, rN az egységnyi idő alatti egyedszám növekedés
A populáció növekedése ideális esetben exponenciális. Ez mindaddig tart amíg, valamilyen korlátozó tényező föl nem lép, vagyis a környezet eltartóképessége „megállásra” nem kényszeríti a populációt. Ha a populáció egyedszáma tehát eléri a környezet eltartóképességének megfelelő egyedszámot (K), akkor a populáció növekedése regulálódik (szabályozódik), a túlélés érdekében.
Megkülönböztetünk r-stratégista fajokat, ahol a reprodukciós rátának megfelelő érték fenntartását célozza meg a populáció és az ún. K-stratégista fajokat. (9. Ábra)
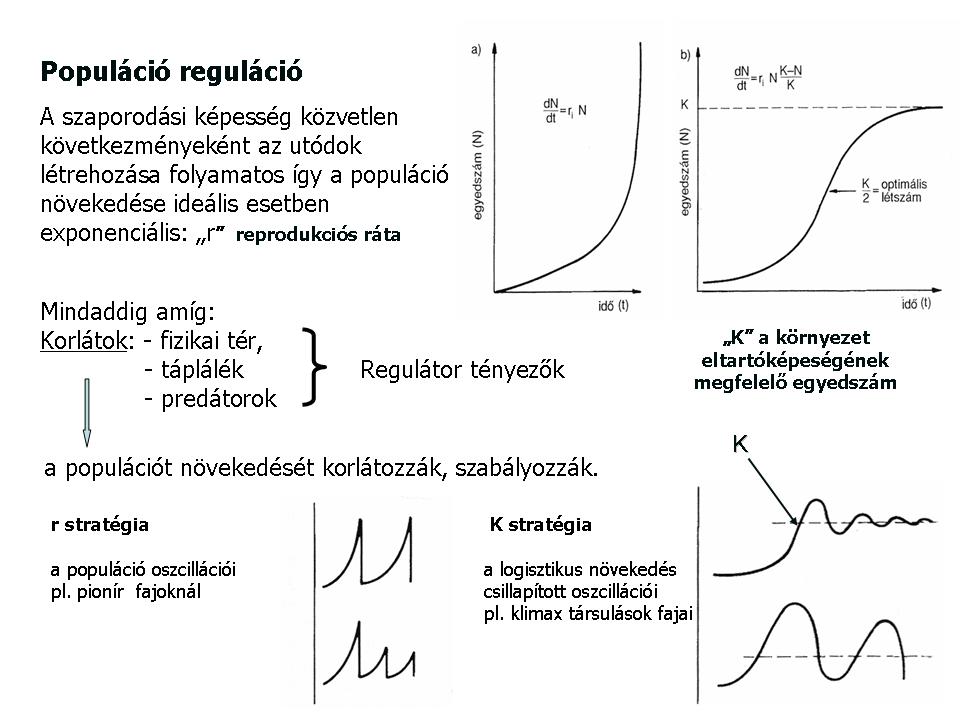
9. Ábra. Populáció reguláció, a K és r stratégiák
Az r stratégia következménye az, hogy a populáció egyedszáma exponenciálisan növekszik majd a növekedés hirtelen ér véget, és a környezet szorító hatására tömeges egyedpusztulás következik be. Így a populáció egyedszáma erősen ingadozó, mert ha a körülmények megengedőek és elegendő forrás áll a populáció rendelkezésére, akkor a populáció gyors egyedszám növekedéssel válaszol, de ha a források kifogynak és a környezeti feltételek leromlanak a mortalitási ráta meredeken újra nőni kezd. Ezt az erőteljes egyedszám ingadozást csak a rövid, vagy egyszerű életciklusú fajok tudják követni. A szántóföldek és kapás kultúrák egyéves gyomnövényei rendszerint r-stratégisták éppúgy, mint egyéb, főképpen pionír élőhelyek fajai. A sok utód (mag) kis túlélési esély a r típusú fajok jellemzője tehát. Ilyen faj például a közönséges tyúkhúr is (10. Ábra). Aszály után a nyárvégi esőzések a késői csírázású gyomflóra gyors megjelenését eredményezi. Ha beköszönt az őszi fagy az egyedek tömegesen pusztulnak el. A jövő évi újbóli sikeres megjelenést a rövid tenyészidőszak alatt létrehozott és talajban felhalmozott bőséges maghozam biztosíthatja.

10. Ábra. Stellaria media – Közönséges tyúkhúr
A K stratégia lényege az, hogy a növekedés a kezdeti exponenciális szakasz után csillapítódik és az idő múlásával egyre kevesebb utód születik. Ha az egyedszám eléri a környezet eltartóképességének határát, vagyis a kritikus egyedszámot (N=K) akkor az utódok létrejötte megáll és a populáció egyedszáma állandósul. Tehát gyors, rövid idő alatti és nagyszámú egyedpusztulás ebben az esetben nem következik be.
A bonyolult és rendszerint hosszú életciklusú fajok követik ezt a stratégiát, pl. erdők fás fajai. Ez a stratégia az utódgondozás feltételén is alapszik.
g) a kompetíciós képesség
A környezeti forrásokért a populációk más populációkkal versengenek. A kompetíció szinte minden élettérben megfigyelhető. Mértéke függ a populációk méretétől, a környezet eltartóképességétől, és a környezet szorító hatásától. A kompetíciós képesség minden populáció sajátja, van, amelyik sikeresebb, van, amelyik kevésbé. Az utóbbi (a „vesztes”), rendszerint kiszorul szélsőségesebb élőhelyekre. A kompetícióban résztvevő populációk ugyanakkor sokféle stratégiát követhetnek.
A kompetíció végbemehet két különböző faj populációja között, ilyenkor interspecifikus kompetícióról beszélünk. Pl. egy erdő különböző fafajainak versengése a fényért, vagy a gyökereik kompetíciója a talajvízért. Fás fajok egyedeinek gyökérzete kompetícióss relációban lehet a lágyszárú fajok gyökereivel is.
Intraspecifikus kompetícióról beszélünk akkor, ha ugyanazon fajhoz tartozó egyedek versengenek egymással. Vagyis szűkös források mellett egy populáció egyedei között is kompetíció léphet föl. Például, gyümölcsösökben a gyümölcsfák sortávolságát azért kell jól meghatározni, hogy az egyedek növekedése ne lassuljon, a termőképesség ne csökkenjen az intraspecifikus kompetíció eredményeképpen. Bármely környezetben az inter- és intraspecifikus versengésnek igen változatos formáival találkozunk. Pl. a madárvilágban a fészekalj fiókái között gyakran intraspecifikus kompetíció érvényesül, ha a szülők nem tudják ellátni őket elegendő táplálékkal. Gyakran a gyengébb fióka elpusztul.
A növényvilágban az intraspecifikus kompetíciónak olyan formái is ismertek, amelyek az állatvilág fajainál nem jellemzők. Ez az infraindividuális versengés (réászletesen lásd. 7. fejezet).
Mint azt már a bevezetőben említettük a növényi populációk testfelépítésének egyéni bélyegei vannak, ezért a növények nemcsak életmódjuk, de testszerveződésük alapján is elválnak az állatvilágtól.
A növényi populációkat ezért specifikus bélyegekkel jellemezhetjük.
A növényi populációk sajátosságai
a) a testfelépítés
A növényi populációk egyedszáma egy olyan specifikum, amely számbevétele csak a növényi testfelépítés megértése után válik értelmezhetővé.
A növények moduláris testfelépítéséből kell újra kiindulnunk. Mint azt már a bevezetőben írtuk, a növények többsége ismétlődő, jól körülhatárolható, elemi részekből épül fel.
A modul vagy elemi hajtás a növényi testnek elemi, szabályosan ismétlődő, morfológiailag jellemezhető strukturális egysége. Egy modul elpusztulhat anélkül, hogy a növényegyed pusztulását okozná, és minden modulnak saját kora és életciklusa van. Ha egy állat például, elveszti bármely végtagját fizikailag súlyosan sérül. Ha egy növény elöregedett és beteg moduljait eltávolítjuk gyakran megújul, megfiatalodik.
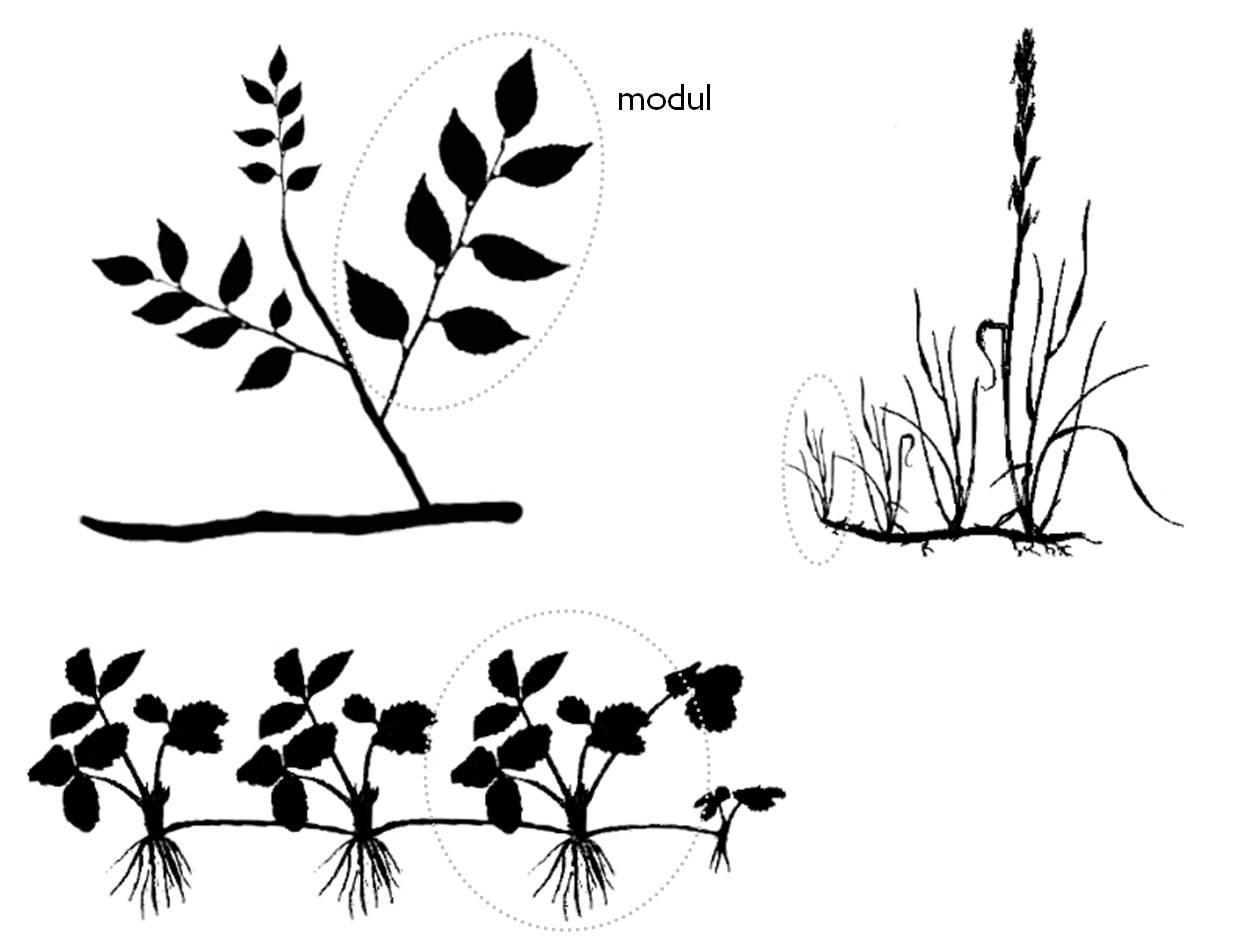
11. Ábra. Fás-, lágyszárú- és klonális fajok moduláris szerkezete
A modulok száma változó lehet, és az élő modulok számát a környezet eltartóképessége is meghatározza. Egy növény a környezeti hatásokra úgy válaszolhat, hogy megváltoztatja moduljainak számát. Ha erős a szorító hatás kevesebb modulból áll az egyed, ha a források bővülnek, akkor több modul fejlődik.
Egy növényi populációban gyakran nem tudjuk, hogy mekkora az egyed és az egyedhatárok is elmosódhatnak (12. Ábra).

12. Ábra. Cornus sanguinea – veresgyűrű som nagyszámú modulból álló példánya (klóntelepe) lösz alapkőzeten
Az egyedszámot meg tudjuk határozni egyrészt úgy, hogy számbavesszük a genetikai egyedszámot, ez a genet (N). Ez azt jelenti, hogy minden egyed egy magból (csírából) fejlődik, tehát a genet a zigóták számával egyenlő. Másrészt a genet mellett fontos lehet (például egy gyökérsarjas cserjénél, vagy vegetatív sarjtelepes egyednél) a modulok száma is. Ekkor számolunk a ramettel (¡). A magvas növényeknél a genet tehát egy mag csírájából fejlődő egyed, melyet a rametek összessége épít föl. Egy genet rametjei tehát mindig azonos génállományúak. (13. Ábra)
A növényi populációk egyedszáma ily módon N¡.
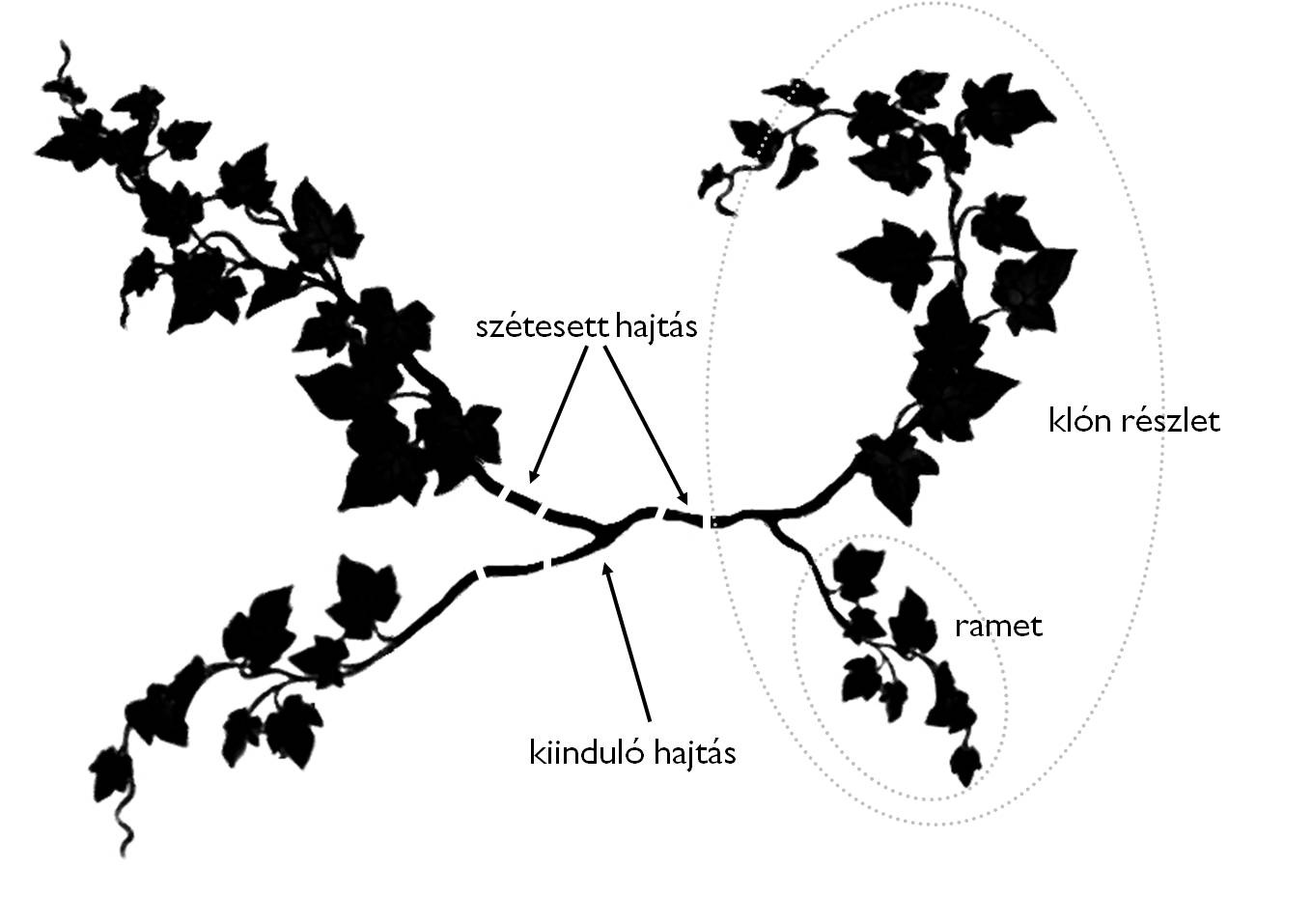
13. Ábra. A genet- ramet szerkezet (van der Maarel ábrája nyomán)
A kertészeti gyakorlatban gyakran a modulok számbavétele is elégséges. Az, hogy mennyi a termőhajtás (termőmodul) egy gyümölcsösben, sokkal fontosabb lehet mint az, hogy hány fát számlálunk, ha azokon nincs egyetlen termőmodul sem (14. Ábra). Szőlőültetvényben a szőlőtőkék száma mellett, a termőmodulok mennyisége is meghatározó. Hiába van sok tőke, ha a modulok száma túl nagy és a kompetíció miatt nem nőnek a fürtök. Ilyenkor ritkítani kell.

14. Ábra : Intenzív almaültetvény: a törpe növekedésű fákon sok termőmodul található
A modulokból vegetatívan új egyedek hozhatók létre pl. dugványképzés, bújtás által. (lásd. vegetatív szaporodásról szóló rész a 4. fejezetben). A vegetatív szaporítás ezen formái a kertészeti gyakorlatban is a növények moduláris szerkezetéből adódnak. Igaz, ezzel csak a rametek száma nő, a genetikai egyedszám nem változik.
Az oltás a modulok összeillesztésének speciális technikája. A fajtanemesítés, szaporítás során gyakran az alanynövényre a nemes fajta modulját (vesszőjét, rügyét) forrasztjuk rá, így megőrizve a fajta előnyös tulajdonságait miközben biztosítjuk azt is, hogy a rezisztens alany életben tartsa. Ha nincsen alany inkompatibilitás, akkor az oltott fajta moduljai gyarapodnak az alany gyökérzetén.
b) Térstruktúra
A növényi egyedek alakját, formáját és a populáció mintázatát jelenti. A magassági növekedés a fény hatására következett be és a modulok elfásodása a fás termet megjelenésével járt. Az oldalirányú növekedés az elágazódáshoz, legyökeresedéshez és a klónszerkezet megjelenéséhez vezetett.
Vannak olyan növényi populációk, amelyeknél mindkét irányú modulképzés erőteljes (15. Ábra).

15. Ábra. Bambuszok (Bambusa sp.) klonális szerkezete a földalatti hajtásrendszer elágazódásából adódik.
A genet-ramet dinamikával a növényi populációk sikeres túlélési stratégiát követhetnek vagy a versengés során előnyösebb helyzetbe kerülnek. Egyes fás gyomok térhódítása a bonyolult térstruktúra kialakításával következik be (16. Ábra).

16. Ábra. Ailanthus altissima - bálványfa gyomosító vegetatív sarjtelepe a Fóti Somlyón. (Udvardy László felvétele 1997-ből.)
Tehát a növények többségénél gyakori a klonális szerkezet, mely az életben maradás és terjedés egyik lehetősége. Klónnak nevezzük az azonos genetikai állományú egyedeket, melyek egyetlen ivaros egyedtől származnak és vegetatívan szaporodva sokasodnak. Tehát a klónok kialakulását genetikai rekombináció nem előzi meg.
A klonális stratégia miatt sok növényfaj igen magas kort is elérhet. Vannak egyes Amerikában élő nyár (Populus tremuloides) egyedek, (genetek), melyek vizsgálata során kiderült hogy koruk eléri 12000 évet, azaz a mag csírázása még a holocén elején következett be. (Mock et al. 2008, Mitton-Grandt 1996)
A változó/változtatható modulszám egy fontos életben maradási stratégia. A környezet szorító hatására változhat az egyed alkotóelemeinek száma. Például egy disznóparéj ( Amaranthus retroflexus) virágzik alig pár centisen is és magot képes hozni néhány modul kifejlesztése után. Ha azonban, a környezeti források lehetővé teszik, méteres magasságig nőhet, majd számos modulja folyamatosan, rengeteg magot érlel és elszórva képes feltölteni a talaj magkészletét több évre is (17. Ábra).

17. Ábra : Szőrős disznóparéj (Amaranthus retroflexus) különböző méretű virágzó egyedei
c) ökológiai életstratégiák
A növényi populációk viselkedését életstratégiákkal jellemezhetjük. A Grime-féle (1979) ökológiai életstratégia összefügg a regulációval is.
Továbbfejlesztve a r, K stratégiát, a versengés mellett Grime az életstratégia típusoknál a környezet szélsőségességét is figyelembe vette.
Három csoportba osztja föl a növényeket
S – stressztoleránsok :
- szélsőséges élőhelyek lakói
- alkalmazkodóképességük rendszerint nagy
- alacsony produktivitás jellemző
Ez az életstratégia jellemző a legtöbb pozsgás növényre pl. kövirózsák (Sempervivum) és varjúháj fajok (Sedum)
C – kompetitorok:
- jó vegetatív terjedésűek
- gyakran klónképzők- kolónia építők
- invazívak lehetnek! pl. Acer negundo (zöld juhar), Cirsium arvense (mezei aszat)
R – ruderálisok:
- egy- és kétéves növényfajok
- nagyszámú magot hoznak, melyek sokáig csíraképesek
- pionír zavart élőhelyek lakói pl. Amaranthus (disznóparéj), Chenopodium (libatop)
Grime átmeneti típusokat is meghatároz, és egyszerű ábrával szemlélteti ezt. A környezet típusa gyakran meghatározhatja azt, hogy egy növényfaj populációjánál melyik stratégia érvényesül erősebben (18. Ábra).
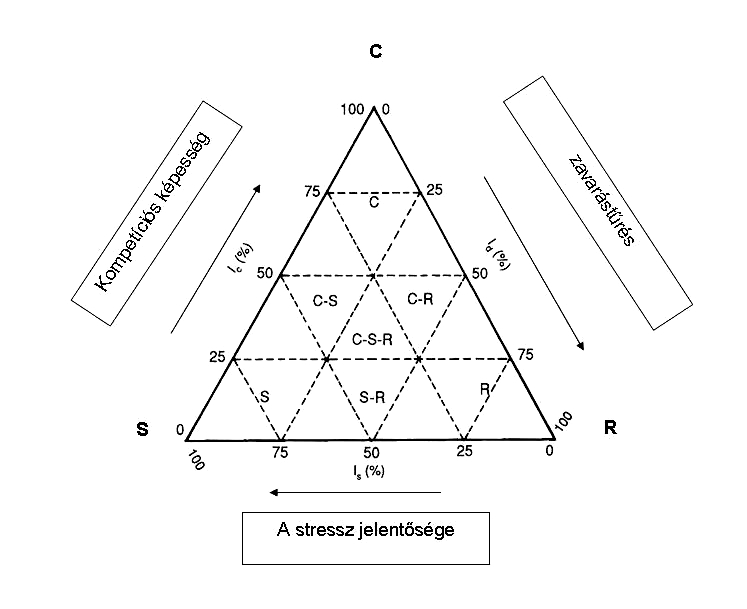
18. Ábra. A Grime-féle életstratégia háromszög
d) rögzítettség
Főképpen a magasabbrendű növények általános tulajdonsága, amely befolyásolja egész életük során a viselkedésüket, a növekedést éppúgy, mint a terjedést.
e) növényi életciklus
John Harper növényi populációökológus a növények életciklusának négyfázisú modelljét szerkesztette meg (1977). Eszerint a növényi egyed életciklusa négy fontos szakaszból áll: mageső, magbank, újulatképzés és egyedi moduláris növekedés. A számunkra látható, utolsó fázis mellett a többi három egyenlő fontosságú a populáció életbenmaradása szempontjából (19. Ábra).
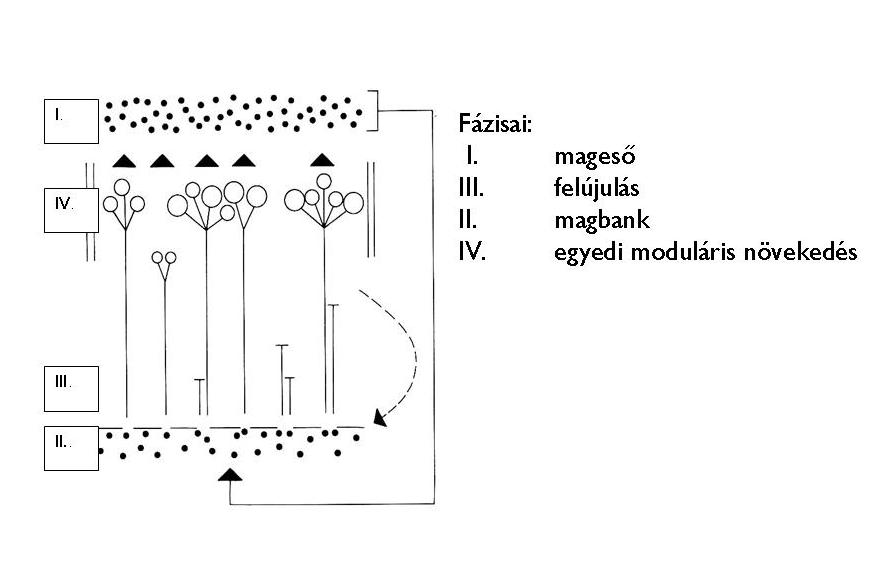
19. Ábra. A növényi életciklus modellje (John Harper alapján)
- Mageső vagy inoculum
A terjesztőképletek – propagulumok – elszórását jelenti. Tágabb értelemben bármely ivaros és ivartalan szaporítóképlet elszórását feltételezi, amely „beoltja” a területet.
Egy populáció magesőjét vagy magára „önti” és ezzel biztosítja túlélését az adott élőhelyen vagy távolabbra szórja, és ezáltal az új élőhelyek meghódítását is biztosítja. Ez a kolonizáció első feltétele.
A mageső (intenzitása) függ a maghozamtól, a mag elszórásának módjától, a magasságtól, a forrástól való távolságtól, a terjedési képességtől, és a terjesztőképlet minőségétől, morfológiai sajátosságaitól (20. Ábra).

20. Ábra. Termések és magok terjesztőképletei. A: Tilia (hárs) – repítő murvalevél; B: Acer (juhar) – ikerlependék; C: Ulmus (szil) – körszárnyas lependék; D: Clematis (iszalag) – repítő(bibe)szálas aszmag; E: Taraxacum (pitypang) – bóbitás kaszat. F: Pinus (fenyő) – repítőszárnyas mag; G: Asclepias (selyemkóró) – repítőszőrös mag; H: Arctium (bojtorján) – kaszatjai horgas fészekpikkelyű fészekvirágzattal terjednek; I: Harpagophytum (ördögcsáklya) – horgas toktermés.
A magesőt befolyásolhatják a környezeti feltételek és az élőhelyi sajátosságok is mint például az időjárás (szélerősség és csapadék), a domborzati formák, legelő és vonuló állatok érkezése stb.
Némelyik növény populációja időzíteni képes a magesőjét. Száraz élőhelyek fajai, parlagok gyomnövényei a csapadékos időszak kezdetén szórják el magjaikat, az állandóan fagyott talajon élő fenyők példaául gyakran tobozaikat csak spontán erdőtüzek idején nyitják föl. Pl ilyen a Pinus contorta Észak-Amerikában.
- Magbank vagy magkészlet
A növények magjai hosszabb-rövidebb ideig nyugalmi állapotban (dormancia) vannak. A nyugalmi állapot kialakulását és megszűnését élettani funkciók szabályozzák, de erősen befolyásolják a környezeti tényezők is. A magok dormancia állapotban terjednek, majd a magbankban halmozódnak föl. Egy élőhely talajában lehetnek olyan magok, amelyek az adott területen élő egyedekkel megegyező fajhoz tartoznak, de más élőhelyről is kerülhet be propagulum. Általában a magkészletben lévő fajszám illetve genetszám nem egyezik a földfeletti populációk fajszámával illetve egyedszámával.
Azt mondjuk a magbank fajlistája a terület potenciális flórája! Egy faj vagy azért hiányzik a területről, mert nincs meg a magkészletben vagy azért, mert nem képes aktiválódni.
Bármely terület diverzitásának értékelésénél fontos számba venni a magbanki tartalékot is. Egy gyommagokban feldúsult, több éve nem művelt talaj a kertész számára nagy veszélyeket tartogat, értéktelenebb lehet.
2. Táblázat: A magbank mérete (magok mennyisége) egyes élőhelyfélességeken
|
Élőhely típusa |
Magszám/m 2 |
|
lucos (kb.30 éves) |
2850 |
|
bükkös |
1000 |
|
trópusi esőerdő |
170 - 900 |
|
mérsékelt égövi legelő |
2000 – 17000 |
|
egyéves gyep |
5400 – 9000 |
|
oroszországi sztyep |
akár 17000 |
|
szántó (búzatábla) |
34000 – 75000 |
|
veteményes (zöldségkultúra) |
1600 - 86000 |
Bizonyos talajlakó orchidea fajok rejtőzködő életmódja is abból fakad, hogy képesek rejtve maradni a talajban (vegetatív propagulumok formájában is), azokban a kedvezőtlen, csapadékszegény években, amikor új maghozamuk kialakítása esélytelen lenne. Ettől még az élőhelyen életképes populációval rendelkeznek.
Egyes növényfajok dormancia állapota nagyon rövid vagy ki sem alakul, ezért ezen fajok a magbankból rendszerint hiányoznak. Pl. fűz (Salix), és nyár (Populus) fajok magjainak nincs dormancia állapota.
A magok életképessége igen különböző lehet, vannak magok melyek néhány évig őrzik meg csíraképességüket, de például egy Tokyo melletti tóban 3000 éves csíraképes lótuszvirág (Nelumbo nucifera) magvakat is találtak. A csíraképesség megőrzése azonban nagymértékben az élőhelyi körülményektől is függ.
- Újulat
A populáció újulat megjelenése az élőhelyen fontos esemény. Nem minden magbankban előforduló propagulumból illetve magból lesz új növény. Egyes magok a talajban elvesztik csírázóképességüket még mielőtt kedvező csírázási feltételek alakulhatnak ki számukra. Másokat a rágcsálók és magfogyasztó populációk tüntetnek el. A gombásodás és más kórokozók terjedése is bekövetkezhet. Végezetül: azok a magok, amelyek ezeket sikeresen megúszták belső élettani szabályozottságuk révén olyan impulzusokat kell kapjanak (akár környezeti hatásra), amely kiváltja a csírázást és az újulat megjelenését. Gyakran az anyapopuláció is képes szabályozni az újulatot. Ha egy erdőben öreg egyedek élnek az újulat sikeres visszaszorítása szinte bizonyos.
Ugyanakkor az ún. gondozó fák – „nurse tree” jelensége is létezik, amikor az újulat fejlődését egy gyorsabban növekvő, akár idősebb egyed segíti azzal, hogy védi őket az erős inszolációtól, szelek szárító hatásától vagy a kártevőktől.
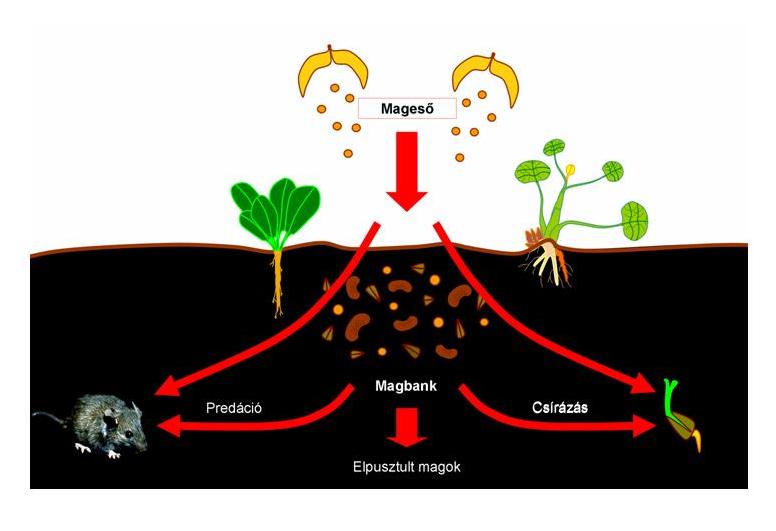
21. Ábra. A magbank sorsa
- Egyedi moduláris növekedés
A növényi populációk egyedeinek csírázása és növekedése előbb-utóbb ahhoz a populáció sűrűséghez vezet el, amikor már a kompetíció korábban említett formái jelentkeznek.

22. Ábra. Különböző számú modulokból álló egyedek növekedése és térstruktúrája
Ellenőrző kérdések
1. Milyen nagyságrendűnek tartják a földi diverzitás értékét fajszámok alapján? A ma élő fajok száma:
- százezres nagyságrendű.
- fél milliós nagyságrendű.
- milliós nagyságrendű.
- 10 millió fölötti.
2. Melyik állatcsoportot (törzset) tartják a legdiverzebbnek a Földön, a fajszám tekintetében?
3. Mekkora az ismert edényes növényfajok száma a Földön?
4. A populáció fogalma.
5. A növényi populációk egyedszámának megítélésében milyen fontos szempontokat kell figyelemben vennünk?
6. Mit nevezünk ökológiai mortalitásnak?
7. A Grime-féle életstratégia típusok közül melyikre illik az alábbi jellemzés?
- jó vegetatív terjedésűek
- gyakran klónképzők
- invazívak lehetnek
8. Soroljon föl 3 tényezőt, ami a mageső sikerét befolyásolja a növényi populációknál.
9. A K és r stratégia lényegek. Egészítse ki az alábbi táblázatot egy-egy-jellemzővel, az első sor mintája alapján!
|
r stratégia |
K stratégia |
|
|
Pl. Magok csírázási üteme |
gyors |
lassú |
|
Növekedési ütem |
||
|
Kompetitív készség |
||
|
Vándorlási készség |
||
|
Az elfoglalt niche mérete |
||
|
Egyedek élettartama |
10. A propagulum fogalma a növényi populációk életében:
Irodalomjegyzék
Borhidi A. 2009: A növényvilág evolúciója és a darwini fejlődéselmélet. Magyar Tudomány. 9.
Cristea, V., Denaeyer, S. 2004: De la biodiversitate la OGM-uri. Ed. Eikon. Cluj- Napoca.
Euforgen adatbázis: http://www.euforgen.org/
Grime, J.P. 1977: Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist. 111:1169-1194.
Harper, J.L. 1977: The Population Biology of Plants. Academic Press.
Hewitt, g. M. 2001: Speciation, hybrid zones and phylogeography, or seeing genes in space and time. Molecular Ecology. 10: 537–549.
Láng I., Bedő Z., Csete L. (szerk.) 2003: Növény, állat, élőhely. Magyar Tudománytár. Kossuth Kiadó. Budapest.
Mock, K. E., Rowe, C. A., Hooten, M. B., DeWoody, J., Hipkins, V. D. 2008: Clonal dynamics in western North American aspen (Populus tremuloides). Molecular Ecology 17: 4827-4844.
Mora, C., Tittensor, D.P., Adl, S., Simpson, A.G.B., Worm, B. 2011: How Many Species Are There on Earth and in the Ocean? PLoS Biol 9(8): e1001127. doi:10.1371/journal.pbio.1001127
Standovár T., Primack, R.B. 1998: A természetvédelmi biológia alapjai. Nemzeti Tankönyvkiadó. Budapest.
van der Maarel, E. 2005: Vegetation ecology. Blackwell Publishing.
Vida G. 1996: Luxus vagy nélkülözhetetlen – Az élőlények sokfélesége. Természetbúvár, 51(4): 10-12.
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028