Plant cell (Citology)
Author: Zsolt Erős-Honti
1.1. The concept of the cell: elaboration and changes of the cell theory
Today, generally accepted is the fact that all living creatures consist of cells. However, till the middle of the 17th century, researchers were not aware that all organisms would be composed of such units. In the 1650’s, Jan Swammerdam, Dutch naturalist observed oval bodies in the blood and later he also discovered that the frog embryo consisted of small orbicles. For the very first time, plant cells were found by an English polyhistor, Robert Hooke in 1663 when examining the cork of woody plants. In his work Micrographia he named the observed structures (resembling the cells of the honeycomb) ‘cellula’ (“cubicle”, “compartment”), thus the denomination of cells themselves also dates back to him. (Although he had thought that the described ‘cellulae’ were small water conducting tubes of the plant, and later it turned out that he only had observed the mere cell walls of dead cells in his microscope, his naming has remained ever since.)
After the first observations, 175 more years passed until the cell concept was generalized to all the known living beings. In 1838, Mathias Jakob Schleiden, a German botanist discussed his observations with Theodor Schwann, a German physiologist in 1838 after that Schleiden determined cellular organization in plants, while Schwann did the same for animals. They published their common observations one year later (1839) in Schwann’s book, where they also gave the points of the classic cell theory:
-
The anatomical, organizational and physiological units of all living creatures is the cell;
-
the cell is a dual entity: it is a distinct, living unit and also the building block of the organism, at the same time;
-
cell assembles from inorganic material (similar to crystals).
According to the last point, Schleiden and Schwann thought that the nucleus of the living cell precipitates and gets outside the protoplasm where later it extends to form new living cells. Consequently, living and non-living stages interchange and thus form a continuity. This can be considered as the survival of the theory of spontaneous generation (‘natura non facit saltus’, i.e. ‘nature does not leap’) accepted since as early as the antiquity. Later, the 3rd point of the classic cell theory was refuted by Robert Remak, Albert Kölliker and Rudolf Virchow who carried out researches on the reproductive processes of the cells. Virchow was the first one to state the fact that ‘all cells come from cells’ (i.e. ‘omnis cellula e cellula’) that is accepted ever since.
1.2. Cell evolution: prokaryotic and eukaryotic cell, different types of the eukaryotic cell
Only indirect evidences do we have on the emergence of the first living cell. According to them, the appearance of the first cell is estimated to have occurred cca. 3.8-4 billion years ago. Though evolution biologists elaborated several theories on the main evolutionary stages of evolving of the first living cells out of the dissolved organic compounds in the ancient oceans, we still have mere assumptions on how the borderline between prebiotic (i.e. the evolution of molecules) and biotic evolution was overstepped.
The structure of the first cells was rather simple. Among recent organisms, bacterial cells have similar composition of such ancient features. The most important peculiarity of these cells is that their genetic material is free in the cytoplasm, and not bound in a real nucleus (i.e. it is not enclose by a nuclear envelope). Due to this characteristic, these organisms are called prokaryotes (‘karyon’ = ‘nucleus’) in contrast with the later (2-3 bya) evolved eukaryotes having an enveloped nucleus containing the DNA. Beside the lack of nucleus, several further distinguishing features can be observed between the two cell types (listed in Table 1): prokaryotic cells lack extended inner membrane system and large organelles; their size is around the magnitude of a micrometre in contrast with the eukaryotic cells of 10-100 microns. The organisation of the DNA in the two cell types is also differing.
Table 1. Comparison of prokaryotic and eukaryotic cell.
|
Prokaryotic cell |
Eukaryotic cell |
|
|
Origin |
3,8 Bya |
2,7 Bya |
|
Size |
~ 1 μm |
10-100 μm |
|
Nucleus |
Missing |
Present |
|
Genetic material |
single circular DNA molecule |
always several, |
|
DNA naked (or associated to non-histon proteins) |
DNA associated to histones |
|
|
1×106 ― 5×106 base pairs |
1,5×107 ― 5×109 base pairs |
|
|
Replication |
Initiated at one single origo |
Initiated at several origos |
|
Cell cycle |
Not observed |
Observed |
|
Cell division |
Fission |
Mitosis, meiosis |
|
Chromosomes are anchored to the cell membrane |
Chromosomes are moved by microtubules of the spindle apparatus |
|
|
Endomembrane system |
Simple |
Complex (ER, Golgi etc.) |
|
Cytoskeleton |
Missing (simple) |
Present |
|
Organelles |
Few |
Always present |
|
Organelles of locomotion |
Protein filaments |
Complex organelles |
|
Ribosome size |
70S (30S + 50S) |
80S (40S + 60S) |
|
Transcription and translation |
In the same compartment |
Spatially separated |
|
Intercellular junctions |
Missing |
Present |
|
Differentiation |
Limited |
Complex |
|
Cytosis |
Missing |
Observable |
|
Multicellularity |
Rarely (e.g. cyanobacteria) |
Usually multicellular organisms |
|
Apoptosis |
Not observed (ambiguous) |
Present |
Despite their basically similar structures, the cells of the different eukaryotic kingdoms (Regnum) may considerably differ from each other (Table 2). The most obvious difference is that animal cells lack cell wall that determines the shape and volume of plant and fungal cells. However, the composition of this organelle is not the same in the later two groups, either: the main polysaccharide of the plant cell wall is the cellulose, while in fungi it is substituted by chitin-like compounds (a molecule basically occurring in animals). Animal cells are usually capable of locomotion: they can move with pseudopodia, cilia or flagella. Though less frequently, but fungal cells sometimes possess flagella (e.g. the zoospores, some gametes or the vegetative cells of the chytrids). Conversely, only the sperm cells of some certain plant groups are motile. (Nevertheless, the sperm cells of angiosperms cannot move actively.) All eukaryotic cells contain smaller or larger vesicles filled with different fluids that are necessary for the transport of previously synthesized or modified macromolecules between the membrane organelles or during the cytotic processes (endo- and exocytosis).
An important derivative of this vesicular system is the lysosomal system in animal cells, where digestive processes and the recycling of degraded material take place within (Quite recently, the presence of lytic vacuoles with a similar role was also proven in plant and fungal cells.) Nevertheless, the main functions of larger vacuoles are space filling and fluid storage in both plants and fungi.
The metabolism of the organisms of the three kingdoms is fundamentally different. Plants are photoautotrophic, i.e. they are capable of producing their own organic compounds from simple inorganic molecules (using light energy). To the contrary, fungi and animals can produce their own organic matter only from other organic compounds, i.e. they are heterotrophic1. The organelles responsible for autotrophy in the plant cells are the chloroplasts that are missing from the other types of the eukaryotic cells.
Table 2. Comparison of plant, fungal and animal cells.
|
Plant cell |
Fungal cell |
Animal cell |
|
|
Metabolism |
autotrophic |
heterotrophic |
heterotrophic |
|
Cell wall |
present, |
present, |
missing |
|
Organelles of locomotion |
cilia, flagella (only on gametes) |
sometimes flagella |
cilia, flagella, pseudopods |
|
Characteristic type of cell cavities |
vacuoles |
vacuoles |
lysosomal system |
|
Specific organelles |
plastids |
Spitzenkörper, lomasome |
centriole |
1.3. Cytoplasm and cytoskeleton
The basic substance of all living cells is the cytoplasm, an aqueous solution of different phases 2. It contains dissolved ions, intermediary metabolic compounds (sugars, amino acids, carbonic acids) and macromolecules (enzymes, nucleic acids). At certain regions (usually close to the cell membrane) the hydration shells of the colloidal particles connect to each other and thus the cytoplasm becomes a jelly-like substance (cytogel).
Enzymes of certain basic metabolic processes are dissolved in the cytoplasm. This is the site of the first stage of monosaccharide degradation (biological oxidation), i.e. glycolysis, as well as that of anaerob fermentation. The most important process of macromolecular metabolism happening here is translation (protein synthesis). It is catalyzed by the ribosomes that are supramolecular complexes consisting of proteins and RNAs (rRNA). The ribosome is composed of a small and a large subunit and its role is to translate the information encoded in the mRNA (i.e. its nucleotide sequence) into the amino acid sequence of proteins (or more precisely, of the polypeptide chain). The role of the small subunit is to bind the mRNA molecule, while the rRNAs of the large subunit catalyze (as catalytic RNAs, called ribozymes) the formation of peptide bond between the amino acids transported to the ribosome by the transfer RNAs (tRNA).
In addition to enzyme proteins, structural proteins are also present dissolved in the cytoplasm. These proteins compose the cytoskeleton ( Figure 1). Cytoskeleton determines the shape of the cells if cell wall is lacking, it is responsible for the constant position and the transport of the organelles within the cell, as well as for the locomotion of the cell. Cytoskeletal proteins are globular proteins polymerized into long fibers. A typical cytoskeletal protein is tubulin that forms the relatively stiff, cylindrical microtubules. A further skeletal protein is the actin, which is best-known from the muscle cells, yet it is present in all eukaryotic cells. To both types of cytoskeletal proteins motor proteins are attached that can move along the fibers (powered by the hydrolysis of ATP).
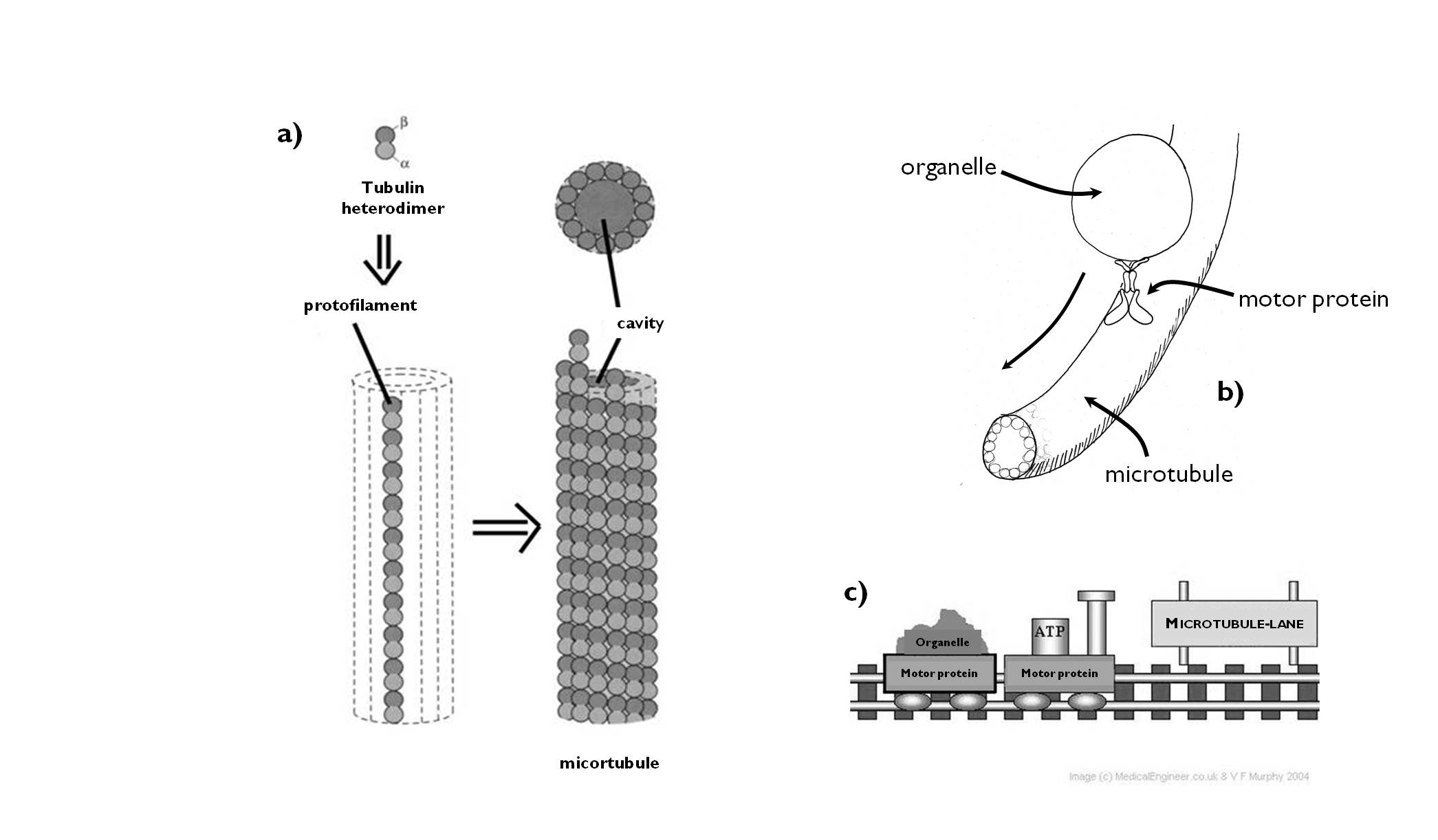
Figure 1. The structure of the microtubules (a) and the concept of motor protein function (b, c).
(a, c: based on internet-derived pictures; a - www.studyblue.com, c - www.medicalengineer.co.uk)
In addition to the transport within the cell, cytoskeleton serves the function of cell locomotion. Both cilia (sing.: cilium) andflagella (sing.: flagellum) are cell projections stiffened by microtubules and totally covered by the plasma membrane ( Figure 2).
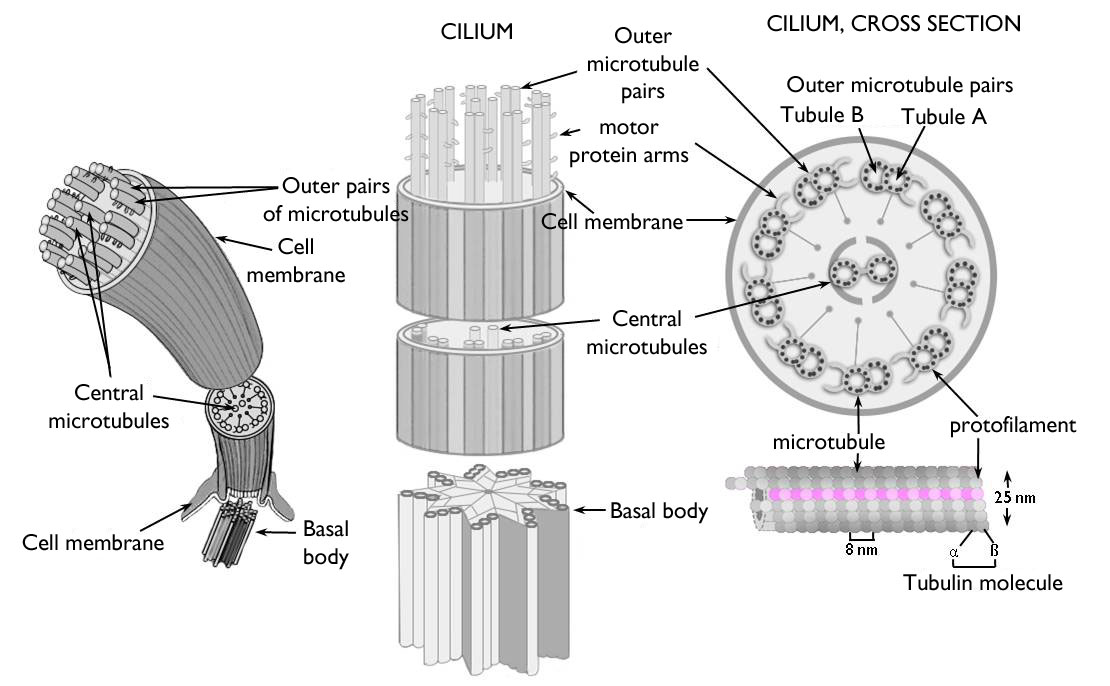
Figure 2. The structure of the eukaryotic cilium (flagellum).
(Based on internet-derived pictures: scienceblogs.com)
1.4. The membrane organelles of the cell
Almost each eukaryotic cell contain a vast endomembrane system. One purpose of this system is to divide the inner space of the cell into compartments of different molecular composition, so as to prevent the interference between the metabolic processes and to separate those metabolic compounds from each other the reaction of which would destroy the cell. (This phenomenon is called compartmentalisation.) The other evolutional advantage of endomembrane system is that it provides a vast inner reaction surface for the biochemical processes catalyzed by the enzymes bound to the membranes.
1.4.1. Cell membrane and transport through the membranes
All living cells are isolated from the environment by a lipid unit membrane, the cell membrane (plasmalemma). It has a double function: on one hand it separates the protoplasm of the cell from the outer environment, but at the same time it plays a significant role in the communication between the cell and the outside world. Special molecules of the cell membrane are responsible for the identical labelling of the cell. Besides, different materials move between the environment and the interior of the cell through the plasmalemma by different transport mechanisms (Figure 3). A further function, characteristic only of the plant cells, is cellulose synthesis carried out by the enzyme complexes (‘rosettes’) embedded in the membrane bilayer.
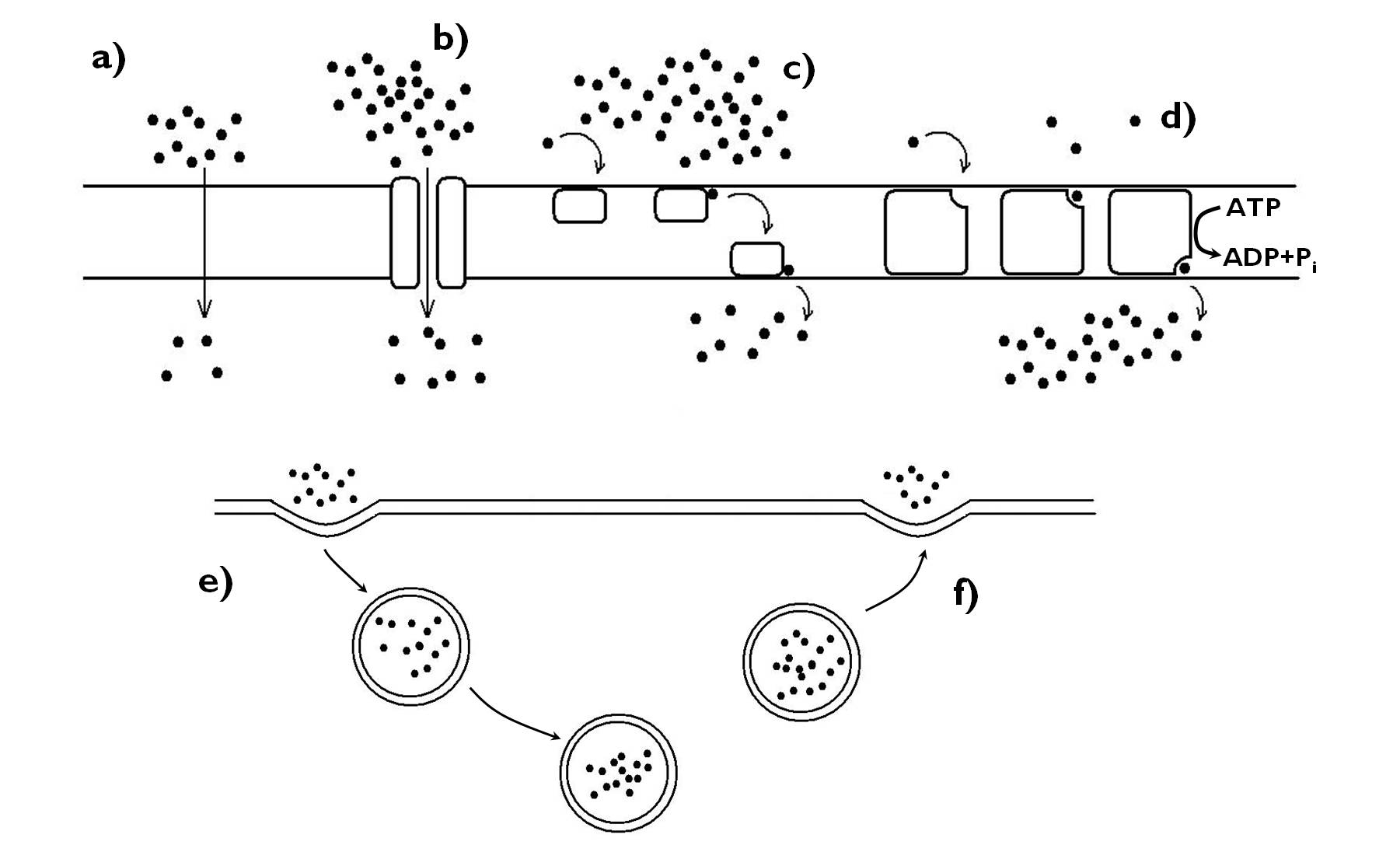
Figure 3. Transport through the membranes. a) diffusion, b-d) protein mediated transport (b: channel protein, c: carrier, d: pump), e)-f) cytosis (e: endocytosis, f: exocytosis).
1.4.2. Secretory apparatus of the cell (ER, Golgi)
An important role of the endomembrane system in the eukaryotic cell is to produce compounds that are released by the cell. The whole process is called ‘secretion’ and the unity of the membrane organelles taking part is the ‘secretion apparatus’ (Figure 4). Secretion apparatus consists of the endoplasmic reticulum (ER), the Golgi apparatus (Golgi bodies), the cell membrane and the membrane bound vesicles transporting the produced compounds between these organelles.
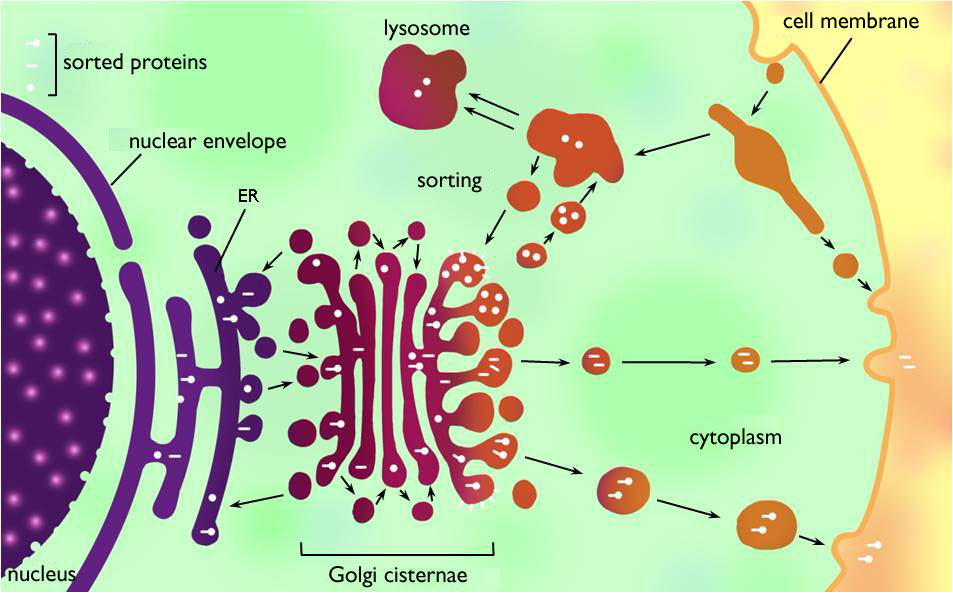
Figure 4. Secretion apparatus and lysosomal system of the eukaryotic cell.
(Based on Kovács)
Endoplasmic reticulum (ER) is the vast, continuous system of interconnected cisternae within the cytoplasm. It is classified into two types: rough endoplasmic reticulum (RER) and smooth endoplasmic reticulum (SER). To the surface of RER ribosomes are attached, this is the reason why it seems to be grained or warted (‘rough’) on electronmicroscopic images. Similarly to those dissolved freely in the cytoplasm, these ribosomes also carry out translation, yet on the surface of RER secreted proteins are synthesized. The proteins produced are translocated into the lumen of the ER during translation (i.e. the proteins are separated from the cytoplasm by the RER membrane) 3. The lumen of the RER is continuous with that of the nuclear membrane surrounding the genetic material of the cell, what is an evidence for the origin of the nuclear membrane from the ER. SER is principally the site of lipid synthesis (e.g. that of sterols and phospholipids) and detoxification, but it also stores calcium ions (if it is necessary within the cell).
Compounds synthesised on the ER get into membrane vesicles bulging out of the ER cisternae and they are transported to the next member of the secretion apparatus, the Golgi body (Golgi apparatus, Golgi complex). Golgi body is composed of several independent membrane sacks (cisternae, dictiosomes) above each other and vesicles serving the transport between the sacks. (However, these sacks are much smaller than the cisternae of ER.) In this organelle, the proteins are modified (chiefly by adding polysaccharid side-chains to them). Further functions of the Golgi are polysaccharide synthesis (but for the cellulose!), the sorting and packaging the macromolecules for the secretion. Modified proteins and polysaccharides exit the Golgi wrapped in secretory vesicles. The vesicles then move to the cell membrane (via the cytoskeleton) where they are finally unloaded by exocytosis.
1.4.3. Vacuoles and crystals
Vacuole s (cell cavities) are characteristic organelles of the plant (and fungal) cell. They are filled with and aqueous solution and surrounded by a unit membrane (tonoplast). The main function of these organelles is to accumulate and isolate such – usually fluid – materials that would interfere with the metabolic processes of the cytoplasm. Such compounds are, for instance, carbonic acids (malic acid, citric acid – often in the form of salts). The fluid content of the vacuole is the cell sap.
The majority of differentiated cells contain few (usually a single), large vacuoles. The reason for this phenomenon is the fact that growth of plant cell is achieved by intense water uptake, what increases the inner pressure (turgor) of the cell that stretches the flexible primary cell wall of the undifferentiated cell and thus enlarges the cell volume. However, this excess water would dilute the cytoplasm to such an extent what would hinder the metabolic processes, so the cell has to transport the water through the tonoplast and isolate it in the vacuole. (Consequently, water also can be regarded as an accumulated material of the vacuole.) Sometimes, accumulated compounds are not fluid but solid ones, e.g. protein crystals – this is one way of the development of aleurone bodies. (Beside vacuoles, storage proteins are synthesised also in the RER.)
The cell sap of the vacuole may contain anthocyans that are pigments of different colours from blue to purple, depending on the pH (e.g. in the epidermal cells of the fleshy scale leaves of red onion or in the similar cells of the red cabbage leaves). Besides, due to their enzyme content, vacuoles are the organelles of hydrolytic processes within the plant cell. (Thus, they perform the function of the lysosomes of the animal cells.)
Since plants – in contradistinction to animals – are incapable of excretory processes, they sequester toxic materials, waste products and excess metabolic products within the cells. A certain way of this sequestration is to turn the harmful compounds (mainly organic acids) into insoluble precipitates. This also achieved in the vacuoles by transporting Ca-ions into the cell sap that here forms insoluble salts with the acids (e.g. CaCO3 /lime, chalk/, Ca(COO)2 /calcium-oxalate/). These salts are present in the vacuoles as various crystal forms (Figure 5). Nevertheless, organic acids themselves can neutralise other toxic materials – e.g. heavy metal ions are bound by maleic acid, citric acid or oxalic acid.
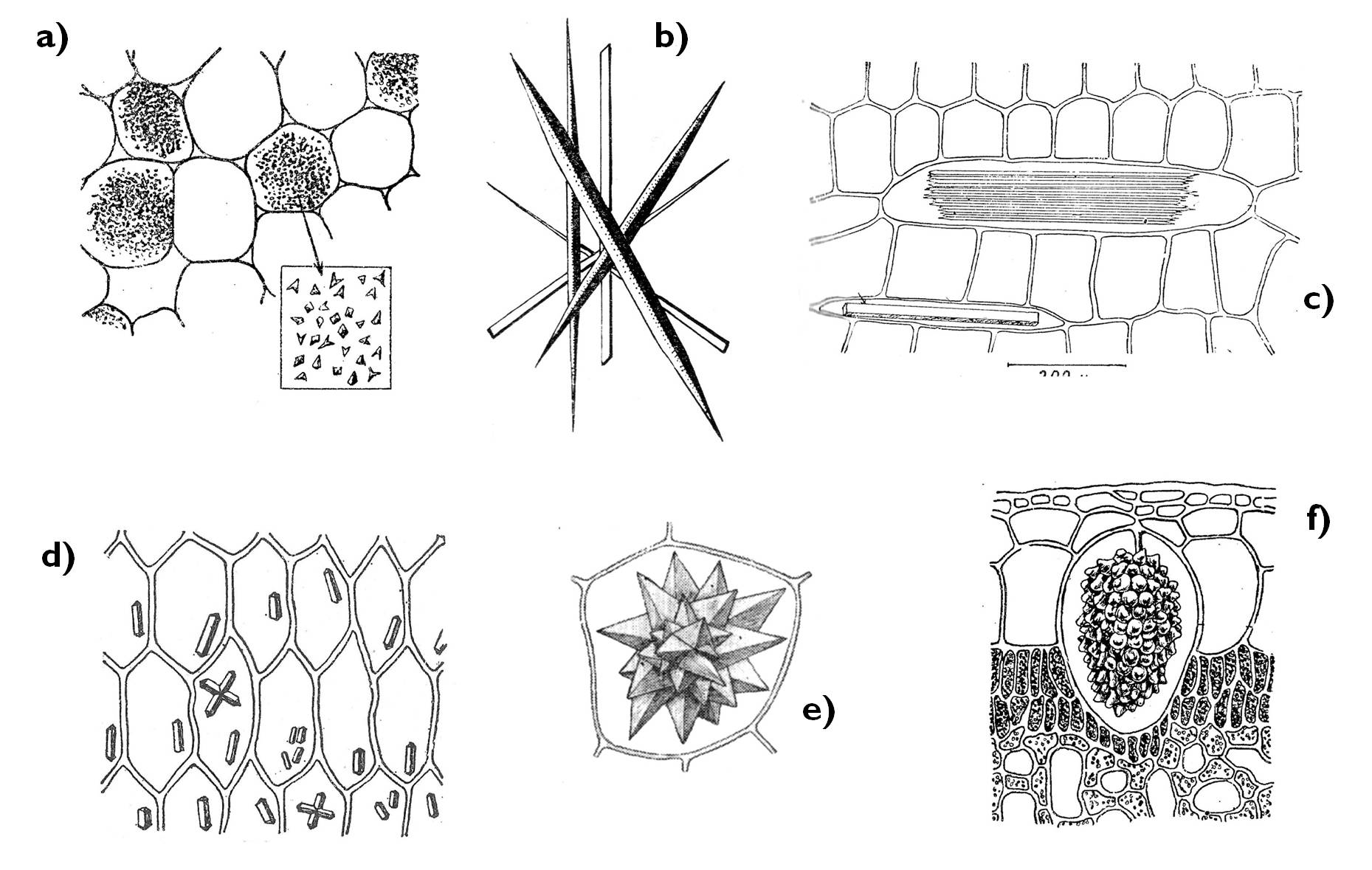
Figure 5. Crystal forms of the plant cell.
a) crystal sand, b) styloids, c) raphide, d) prismatic crystals, e) druse (rosette crystal), f) cystolith.
(Based on Sárkány and Szalai)
1.5. Cell wall
To the outside from the cell membrane, the majority of cells are surrounded by a more-or-less stiff, rigid organelle, the cell wall. The cell walls of bacteria, algae, fungi and plants have basically similar function and structure (though the composing molecules vary, according to the different taxonomic groups). Quite rare are those plant cells that lack cell wall. These are either surrounded by other cells with walls or they live for only a short time (e.g. gametes). An exceptional case is that of the artificially elaborated protoplasts. From these plant cells of different model organisms of molecular breeding researches the cell wall is removed by an enzymatic treatment.
The main function of cell wall is defence. It protects the cell from the outer physical impacts and also against some chemical compounds, but it may inhibit the penetration of parasites or impede the chewing of herbivorous insects, as well. A further important role is to prevent the harm caused by the intense turgor pressure in case of increased water uptake (e.g. in intensely growing cells). Due to its relative rigidity, it determines the shape and size (volume) of the plant cell. Nevertheless, it is worth mentioning that cell wall is a plastic organelle, it can react to the changes of the environment or to the impacts of the protoplasm: due to the enzymes present, the cell wall is under continuous remodelling. This is the reason why we cannot regard it as a non-living part of the cell.
In certain storage organs (e.g. in the endosperm of some seeds) the polysaccharides of the thick cell wall serve as accumulated materials that can be mobilized later by the degradation of the wall. Thus, cell wall may be a storage organelle of the cell, too.
1.5.1. Formation of plant cell wall
In the thelophase of cell division (mitosis or meiosis, see below in details) the protoplasm of the mother cell is divided in the process of cytokinesis to form two daughter cells. At the same time, perpendicular to the original cell walls a new proportion of wall is formed. (Thus, the majority of the cell wall of the daughter cells is basically identical with that of the mother cell, and new wall is synthesised only on the edge surface between the progenies.)
Under high magnification, cell wall has a stratified structure. The neighbouring cells are attached to each other via the middle lamella, a pectin-rich layer between their cell walls. Right beneath the middle lamella is the primary cell wall, which is the first synthesised form of the wall after the cytokinesis. Primary wall is thin, plastic and still expandable; this is the only cell wall layer of undifferentiated (e.g. meristematic) cells. Later, during the process of differentiation – usually three – further layers are juxtaposed onto the primary cell wall. These comprise the secondary cell wall that is much more thick, rigid and incapable of further expansion.
The precursor of the cell wall (cell plate) produced in the telophase is not totally continuous: scattered pores are to be observed on it, which enclose ER-cisternae. These pores remain in the cell wall during the formation of the cell wall layers, and they serve the cytoplasmic continuity between the neighbouring cells. These protoplasmic bridges are called plasmodesmata (sing. plasmodesma).
1.5.2. Composition of the cell wall
The general composition of the cell wall is rather similar in case of the different taxonomic groups: a main skeletal polysaccharide (murein in bacteria, chitin-like compounds in fungi and cellulose in the plant cells) stabilized by hydrogen bonds is embedded in a matrix consisting of other polysaccharides, proteins and further special compounds. The molecules of the matrix are attached to the skeletal compound also via hydrogen bonds. Beside structural molecules, cell wall contains enzyme proteins, as well, that are responsible for the elongation and other changes of the cell wall structure, and also for its role in defence against pathogens.
The framework of the plant cell wall is composed of the linear chains of ß-linked-D-glucan, namely cellulose. Cellulose chains arranged parallel are interconnected by strong hydrogen bridges, what results a strictly ordered, crystal-like structure. (These ordered regions – called micellae – are connected to each other by less-ordered ‘paracrystalline’ regions, hardened by less hydrogen bond.)
The ground substance of the cell wall is the matrix. This partially consists of different polysaccharides. One of these is the hemicellulose that is a general term for heterogeneous short-chained polisaccharides composed of various monosaccharides (glucose, arabinose, xylose, mannose etc.). They are attached to the cellulose microfibrils via hydrogen bonds, and they enwrap them. The entire cell wall matrix is impregnated with pectins, which are polysaccharide compounds, too. Their main purpose is to glue the other molecules of the cell wall to each other. (This very same function accounts for the fact that middle lamella, sticking the neighbouring cells to each other, is almost entirely composed of pectins.) Both hemicelluloses and pectins are produced within the Golgi apparatus and gets into the cell wall by exocytosis.
In addition to polysaccharides, proteins with sugar chains (glycoproteids) are also present in the cell wall. Some of these are enzymes taking part in cell wall elongation (e.g. expansin, hydrolases, peroxidases or transferases). Others are structural proteins that support the wall matrix by binding to the polysaccharide elements via hydrogen bonds (e.g. the extensins).
All the up-to-now mentioned components (cellulose, hemicelluloses, pectins and proteins) are present in both the primary and the secondary cell wall. On one hand, the considerable thickness of the secondary wall is due to the fact that it contains a larger amount of these materials. The common molecules are just slightly modified: the proportion of the compounds somewhat changes, as well as the orientation of the cellulose microfibrils (they run irregularly in the primary wall, yet they are helically oriented in the secondary wall).
Beside the constituents of the primary wall, further characteristic compounds are built into the secondary wall. The most important of these are lignins. Lignins are phenolic polymers (composed of coumaril alcohol, syringyl alcohol, guaiacyl alcohol etc.) with a composition being characteristic of the taxonomic position. The cross-linked polymer entwines all the secondary wall and confers mechanical strength to it (similarly to the steel skeleton of the armoured concrete). Lignified cell walls isolate the protoplasm of the cell from the outer environment to such an extent that causes the death of the cell due to the limited nutrient uptake. This phenomenon is clearly observed during the differentiation of the xylem in the vascular bundles or in case of sclerenchymatic fibres.
1.5.3. Plasmodesmata
Water in the plant body occurs in two different spaces, in the apoplast and the symplast. Apoplast means all the water outside the protoplasts (i.e. the plasma membranes) of the cells, i.e. the cell wall continuum together with intercellular spaces. To the contrary, the interconnected protoplast of the cells is called symplast. Due to the characteristic contact of the cell walls of the adjacent cell obvious is the fact that the apoplast is a more-or-less continuous system within the plant. (An exception is the apoplast of the root cortex and that of the root stele, since the suberinised cell walls in the endodermis isolate the two system from each other. – In details see in Chapter 3!)
Though not so obviously, but the symplasts of the neighbouring cells are also interconnected – similarly to the apoplast – so the plant body represent an almost complete symplastic system4. The protoplasts of adjacent cells are connected to each other via cytoplasmic bridges called plasmodesmata.
1.5.4. Cell wall thickening
In the majority of differentiated plant cells, secondary cell wall layers are deposited onto the inner surface of the primary wall. In these cases, the cell itself produces the compounds of the secondary wall and the cell lumen decreases during the progress of thickening. This is centripetal cell wall thickening (e.g. in case of tracheary elements). Less frequently, secondary cell wall compounds are synthesised by other cells surrounding the differentiating one – for instance in case of spores or pollen grains. Consequently, new layers are laid onto the outer surface of the primary wall, and so the size of the cell lumen does not alter. This process is called centrifugal cell wall thickening ( Figure 6).
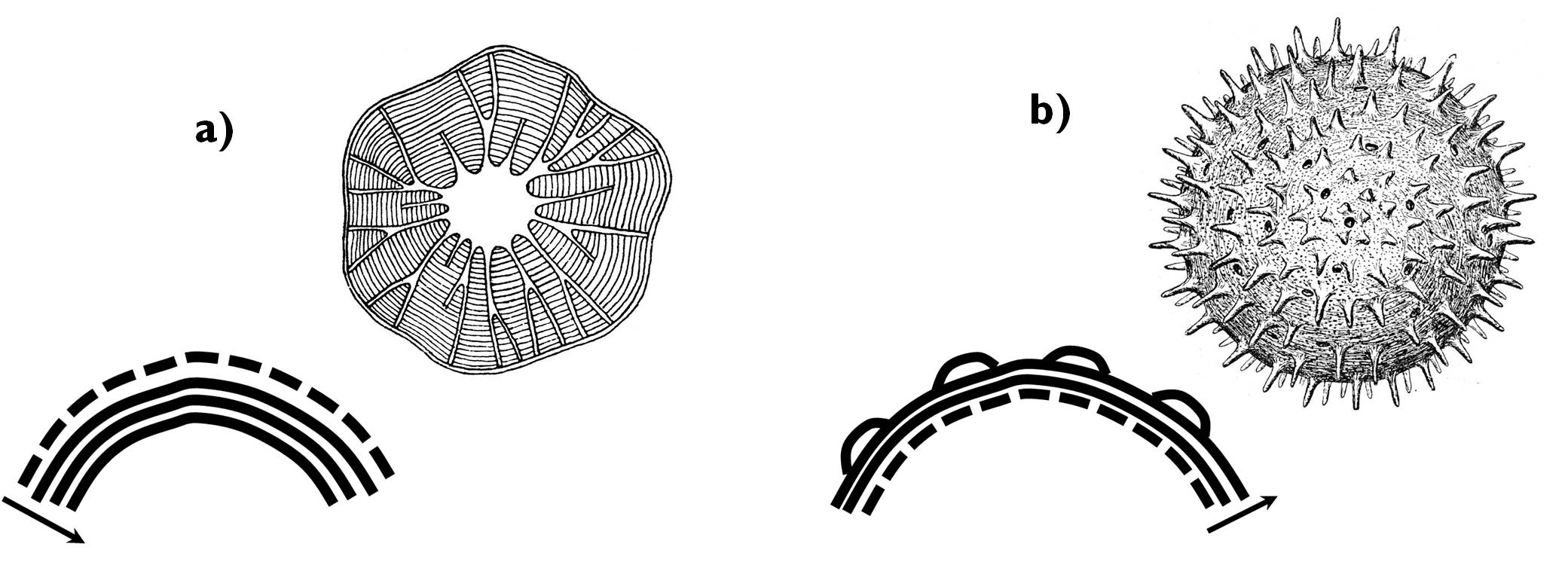
Figure 6. Comparison of centripetal (a) and centrifugal (b) cell wall thickening.
(Dashed line: primary cell wall, continuous lines: layers of the secondary wall.)
(Based on: a - Kárpáti; b - Sárkány and Szalai)
1.6. Mitochondrion and plastid: the organelles of energy production
All living cell needs energy to maintain its vital processes. This energy is available for the enzymes of metabolic reactions in the form of the high-energy pyrophosphate bond of the molecule adenosine-triphosphate (ATP). Within the eukaryotic cell, intense ATP synthesis occurs inside the mitochondrions and the chloroplasts (and with a much less intensity in the cytoplasm). From the mitochondria, the majority of the ATP molecules are transported into the cytoplasm where they are utilised in other energy consuming processes. To the contrary, ATP synthesised in the chloroplasts remains in the organelle and as the storage molecule of light energy it is used for producing organic compounds from inorganic molecules (i.e. for the Calvin-cycle).
1.6.1. Evolutionary origin of the mitochondrion and the plastid
As described above, in the chapter on the evolution of the eukaryotic cell, the origin of the mitochondrion and the plastid is interpreted by the endosymbiotic theory of Lynn Margulis. Accordingly, mitochondrion and plastid are supposed to have been free-living independent prokaryotic cells: the ancestor of the previous belonged to the group of the purple non-sulphur bacteria capable of efficient aerobic catabolism (tricarbonic-acid-cycle, terminal oxidation), while plastids derive from ancient photosynthetic cyanobacetria.
As a result of their similar evolutionary origin, mitochondria and plastids share several common structural and even functional characteristics. Both organelles are surrounded by a double membrane: a tense, smaller outer membrane and an expanded inner membrane. The latter folds into the ground substance of the organelle and bears several different enzyme molecules embedded within. Similar is the background of the ATP synthesis in the two organelles as being based on the unequal H+-concentration on the two sides of the inner membrane.
1.6.2. Mitochondria: structure and function
Mitochondria (sing. mitochondrion) are oval or filiform organelles up to some micrometres, present in all eukaryotic cells capable of aerobic catabolism (i.e. cellular respiration). The number of mitochondria varies according to the energy demand of the cell. The inner membrane covering the organelle forms vast infoldings called christae (Figure 7). (Beside christae containing mitochondria, tubular ones also occur, e.g. in the cells of certain fungal species.) The ground substance of the mitochondrion enclosed by the inner membrane is the matrix. This region contains the circular DNA molecules (almost always in several copies) bearing the genom of the organelle, as well as the particles and molecules required for protein synthesis (ribosomes, tRNAs, mRNAs).
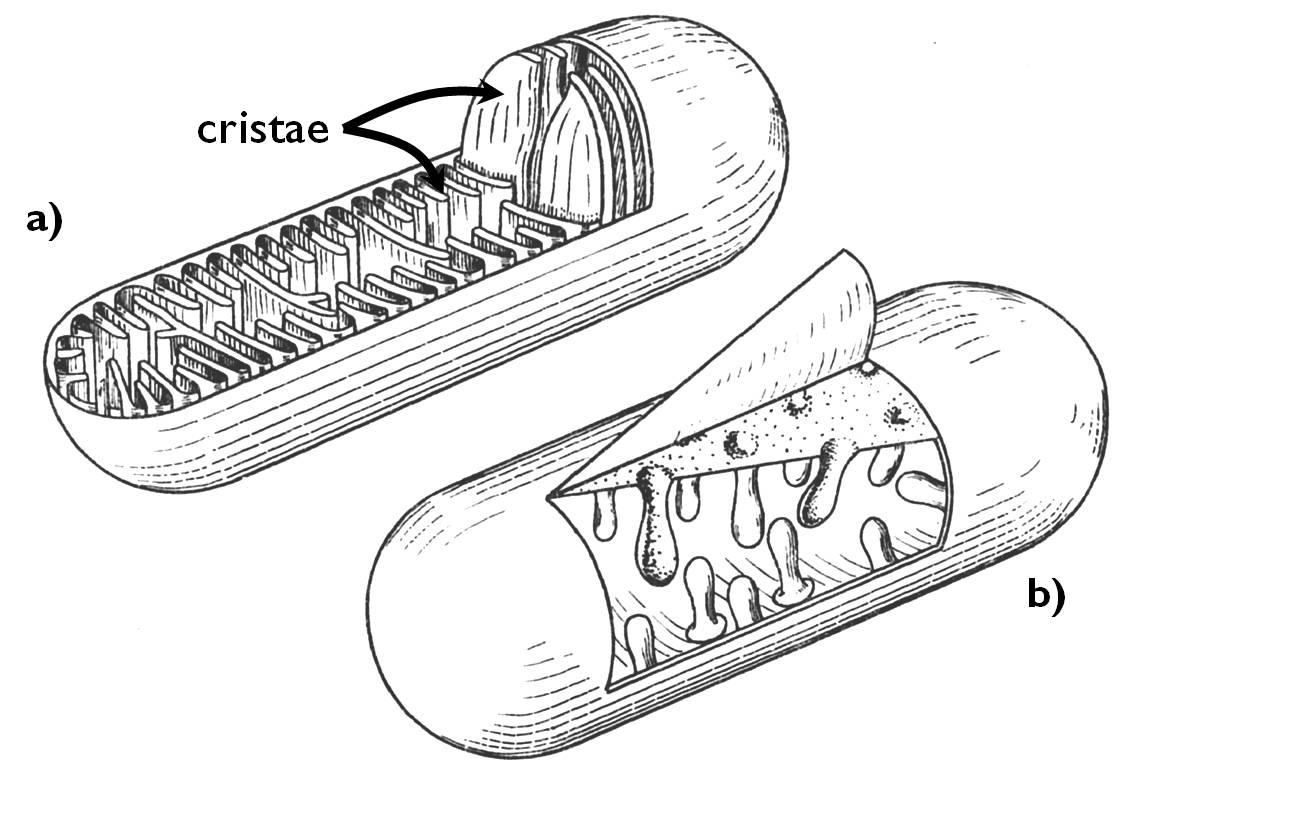
Figure 7. The structure of a mitochondrion with cristae (a) and tubules (b).
(Based on Haraszty)
Mitochondrion is the organelle of cellular respiration. Within its matrix takes place the citric-acid-cycle while the process of terminal oxidation is carried out by the electron transport chain within the inner membrane.
1.6.3. Plastids: structure and function
Plastids are organelles of various shape and size. Their number is also variable within the plant cell, some algae contain one single plastid, yet an average photosynthesising plant cell bears 20-60 pieces of them. General features of plastids are their double membrane envelope and the fact that inner membrane comprises a highly folded inner system of cisternae called tylakoid. The ground material of these organelles is the stroma. Similarly to the matrix of the mitochondrion, stroma contains the own ring-shaped DNA, different RNAs and ribosomes (like those of the prokaryotes) responsible for protein synthesis.
1.6.3.1. Chloroplast
Chloroplasts conduct the photosynthesis, the process that determines the metabolic particularity (photo-auototrophy) of green plants. Its scientific name (‘chloros’ = ‘green’, ‘plastos’ = ‘moulded’, ‘modelled’) derive from its characteristic colour that is caused by the chlorophyll pigments playing a crucial role in photosynthesis. Chloroplasts occur in the cells of all green tissues of the plant (in the chlorenchyma of leaves, young stems, unripe fruits etc.). An ultrastructural feature of the chloroplast is that their tylakoid membrane is arranged in stacks called grana (sing. granum) ( Figure 8).
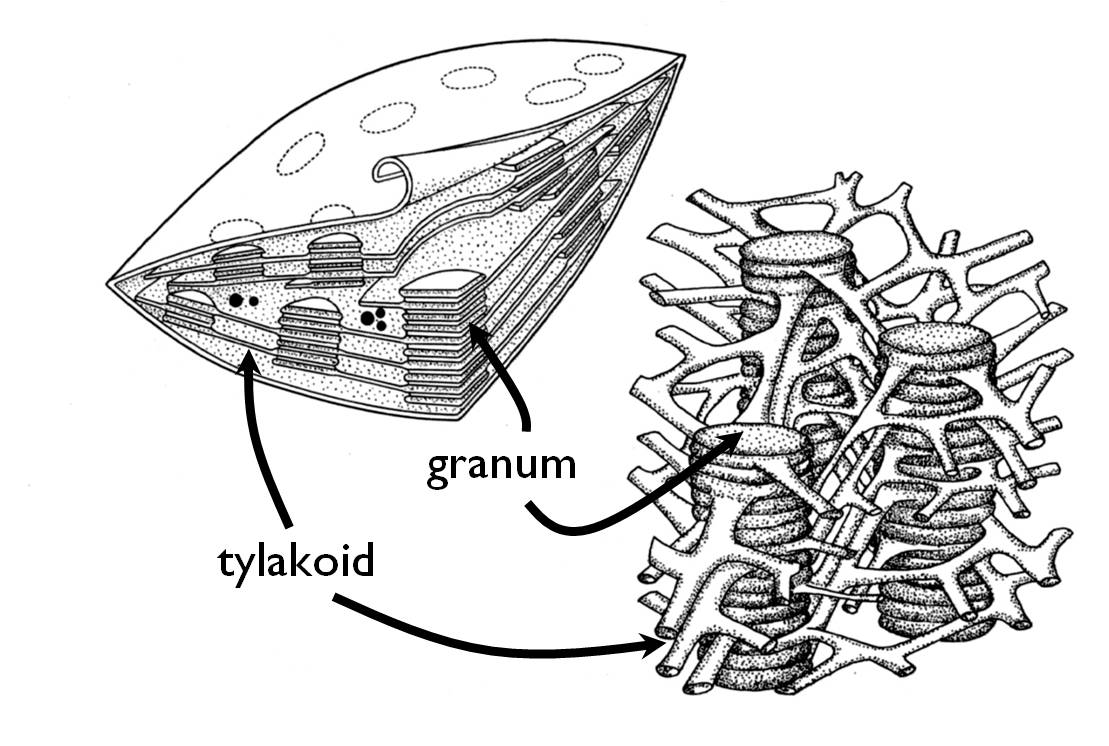
Figure 8. The structure of the chloroplast.
(Based on Haraszty)
In the process of photosynthesis inorganic compounds (i.e. water and CO2) are converted into organic matter (glucose) using light as a source of energy. At the same time, oxygen is released as a by-product. Photosynthesis occurs in two stages. In light dependent reaction solar energy is converted into chemical energy (ATP), while water molecules are degraded into O2 and H+-ions and electrons. The latter two finally get to NADP molecules forming NADPH+H+. The aim of light independent reaction is to reduce CO2 to glucose with the hydrogens transported by the NADPH+H+ and the energy of the ATP. The two stages can be precisely localized within the chloroplast: light dependent reaction is catalyzed by the enzymes dissolved in the stroma, while the light independent reaction happens attached to the tylakoid membrane.
Glucose released in the chloroplast is either transported out to the cytoplasm (in the form of triose-phosphates) or it is temporarily accumulated in the chloroplast. In the latter case it is transformed into starch, thus in functioning chloroplasts starch grains are to be observed periodically 5.
1.6.3.2. Further types of plastids
Plastids of the meristematic cells are undifferentiated; they contain rudimentary tylakoid membrane without grana. These plastids of such simple structure are the proplastids that can develop into any other plastid type. Proplastids are also capable of division by binary fission.
Storage organelles are the different forms of leucoplasts (Figure 9). These may accumulate proteins (proteinoplasts), oil (elaioplasts) and most often starch (amiloplasts). In case of each type, accumulated substances are surrounded by two membranes (similarly to the envelope of the chloroplast) and their tylakoid system deteriorates. Proteins are stored e.g. in the plastids of the epidermal cells of Commelinaceae (e.g. Zebrina pendula), elaioplastids are most frequently observed in seeds and fruits.
A further type of plastids is the chromoplast. These contain so high amount of carotenoids that these molecules precipitates as crystals within the plastid and thus they distort the shape of the organelle according to their crystal form. Cromoplasts are responsible for the reddish orange colour of some plant structures (storage organs, fruits, flowers) (Figure 9).
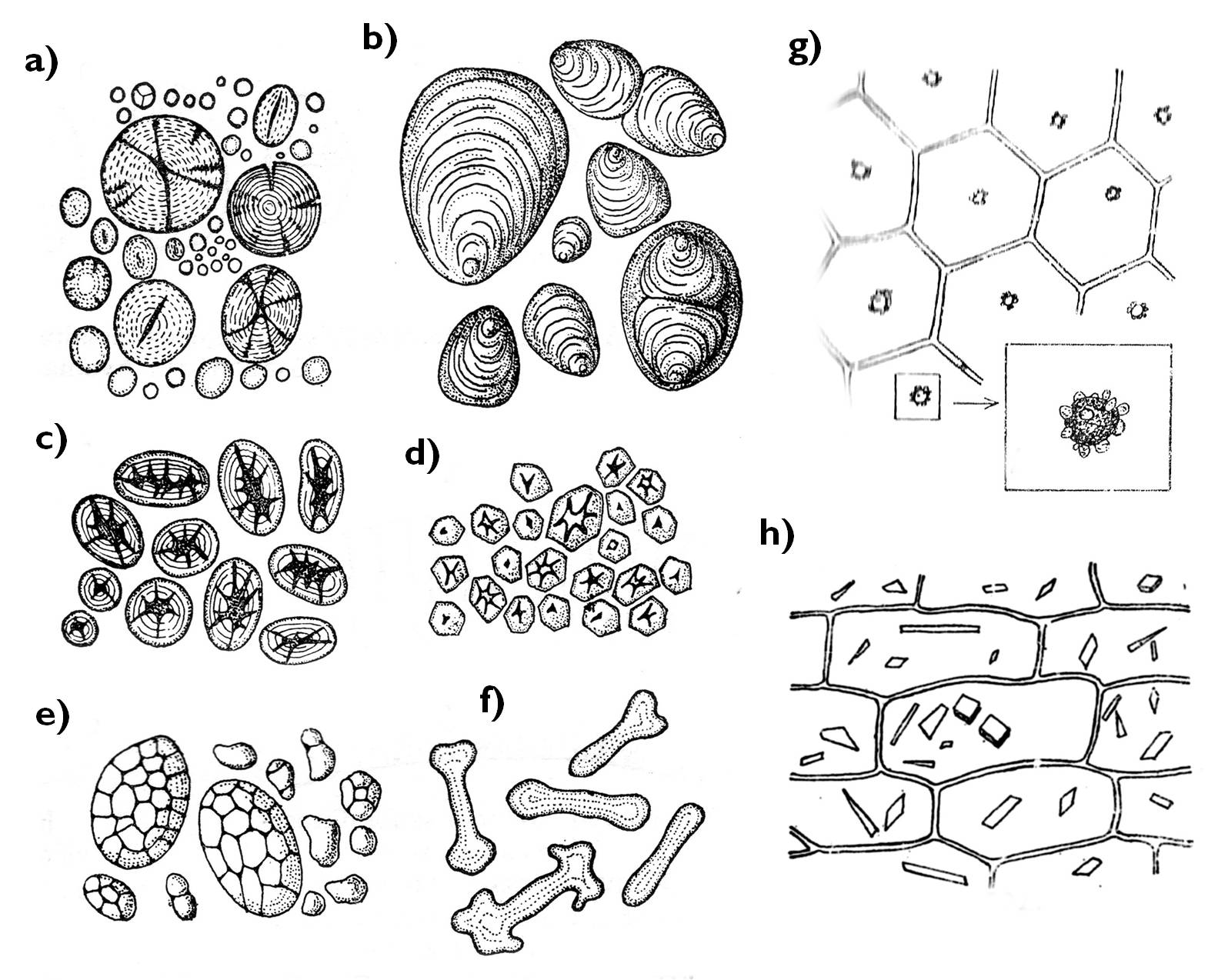
Figure 9. Types of plastids. a)-f) amiloplasts (a: Triticum, b: Solanum, c: Phaseolus, d: Zea, e: Avena, f: Euphorbia); g: proteinoplast (Zebrina), h: chromoplast (Daucus).
(Based on : a-f: Kárpáti , g-h: Sárkány and Szalai )
1.8. Nucleus: structure and function
1.8.1. The structure of the nucleus
The most characteristic organelle of the eukaryotic cell that distinguishes it from the prokaryotic ones is the nucleus. The genetic material of the previous cell type is not present free in the cytoplasm, yet is surrounded by a double unit membrane (nuclear envelope). Nuclear envelope mot probably evolved from the cisternae of the RER. The close relationship between these organelles is obvious even today: the membrane of the RER is continuous with that of the nuclear envelope. Moreover, the structure of the organelle is quite similar: ribosomes are attached to both, and the lipid composition of the membranes is also alike.
1.8.2. Organisation of the genetic material
The double helix of the DNA is a rather thin complex, thus it is a very vulnerable and fragile molecule. Since the information is coded in the DNA in the form of the nucleotide sequence (similarly to the sequence of letters coding the information of the written text), all structural damage of the DNA cause some loss of information. As prevention against such damages, DNA does not float free in the nucleus, yet it is spooled around histone proteins ( Figure 10). The structural unit composing of the DNA and the histones is called nucleosome. In the nucleus, genetic material is always present in the form of nucleosomes, i.e. DNA is never unwound from the histones. Even polymerize enzymes moves around the histone discs during transcription.
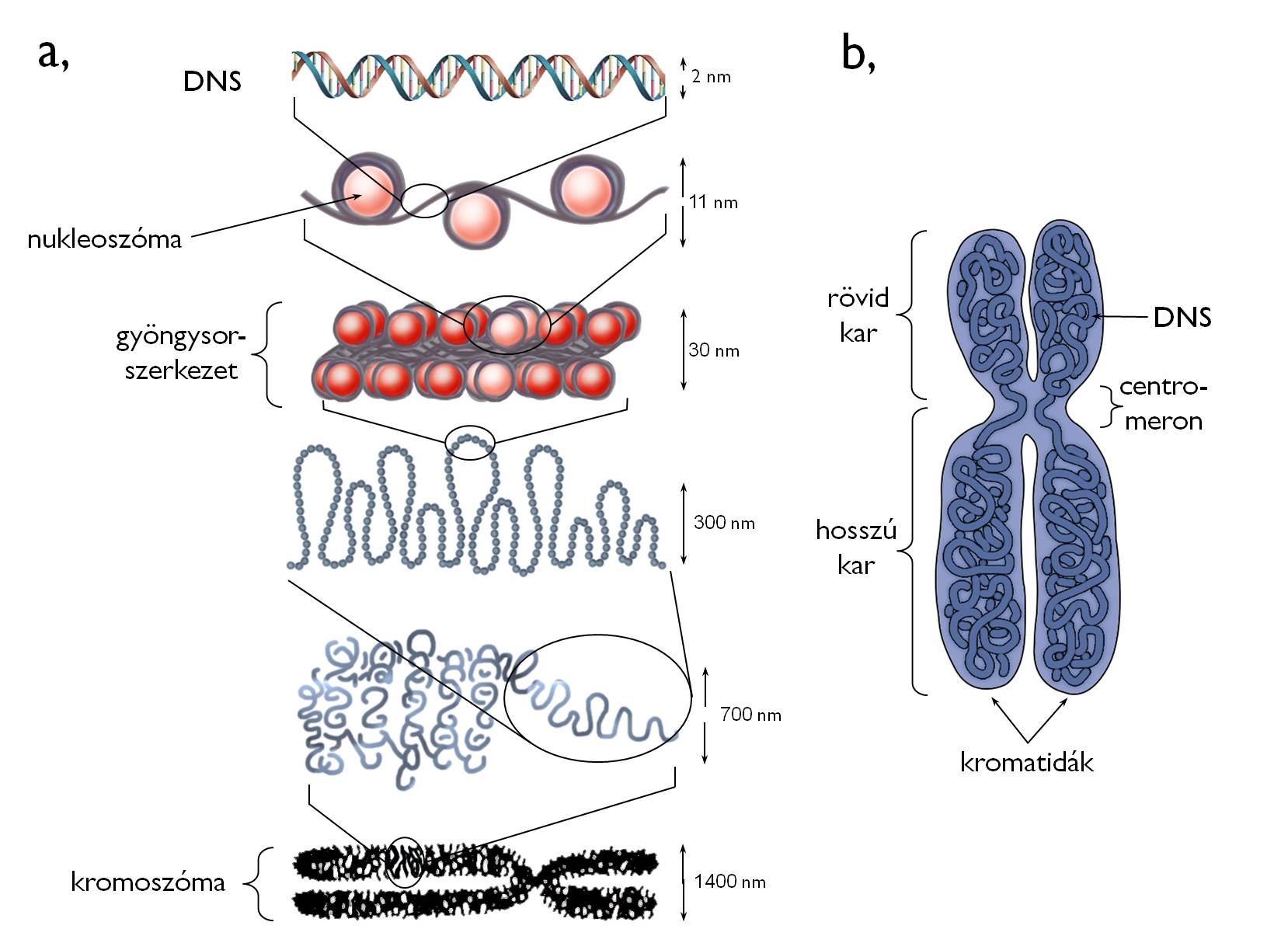
Figure 10. The organization of genetic material (a)
and the structure of a chromosome (b).
(Based on internet-derived sources: a - biology.westfield.ma.edu, b - www.downsyndromenipt.info)
The most densely packed form of DNA is the chromosome, observable during cell division 6. In the chromosomes, DNA wrapped around the histone discs is attached to the scaffold proteins serving as a framework of the chromosome. The most important advantage of forming chromosomes is that during the division DNA can be transported and sorted between the daughter cells with the minimum risk of structural damage.
Chromosomes are usually composed of two halves called chromatids (Figure 10). (Even between the two cell divisions, the sister chromatids of a certain chromosome are in close vicinity. However, in this life stage of the cell chromosomes may contain one single chromatid. – See below in details!) Chromatids are attached to each other at the centromere (primary constriction). (Besides, secondary and tertiary constrictions may also be presented on the chromatids.) Centromeron dissects each chromatid into two arms. At the end region of the arms telomers are found. These regions of highly repetitive sequences protect the coding DNA segments (i.e. the genes and regulatory regions) from deterioration causing the loss of information.
Each chromatid contains a single DNA double helix. If two sister chromatids comprise a chromosome, it contains two DNA molecules. The nucleotide sequence (and hence the coded information) of these two DNAs is completely identical. The reason is the fact that sister chromatids are born during the replication as the two identical descendants of the previously single DNA.
Chromosomes contain the genes. Gene is a certain sequence of the DNA containing the information responsible for a single feature (or more precisely for a protein or an RNA molecule). The region of the chromosome where a certain gene is located is called locus. A particular character (e.g. shape of the pollen or the flower colour) usually have several varieties among the individuals of the species (some plants have elongated others round pollens, white or red flowers). In these cases the genes responsible for the traits also have different forms called alleles7. The entirety of all the genes of the cell (i.e. of the organism) is the genome. It is important to emphasise that only a part of the eukaryotic genome is present in the nucleus (nuclear genome), because some other organelles (mitochondrion, plastid) also contain DNA (extranuclear genome) 8.
Consequently, at a certain locus both chromatids of a chromosome inevitably bear the same allele of a given gene (because of the identity of the DNA sequences). Genes of a chromosome compose a linkage group, what means that (apart from the case of chromosomal damages) a certain chromosome has the same set of genes in each member of the species i.e. the alleles of the same set of features (though these alleles may be different).
In the vegetative cells of the plant body, each gene is present in two copies, since in the nucleus of these cells contain two copies of each chromosome called homologous chromosomes (homologs or homologues). Homologous chromosomes carry the same types of genetic information (i.e. the same genes), yet not necessarily in identical forms (i.e. not the same alleles). The difference between the homologues is their origin: one is inherited from the egg cell and the other from the sperm cell. Some cells contain only one of the homologues – these are called haploid cells. In diploid cells both members of the homologous chromosomes are present. Gametes are haploid cells in each living organism, and the zygote produced by fertilisation is a diploid cell. In plants, some cells contain three, four or even more copies of a given chromosome (i.e. the genes of this linkage group). These are called triploid, tetraploid or in general term poliploid cells.
1.8.4. Cell division
During the development of the plant body, as well as within the life cycle of the plant, cells have to multiply themselves. A necessary part of this process is the share of genetic information (i.e. the DNA) between the daughter cells, what requires the dense package of the fragile DNA molecule into chromosomes. Cells entering the division always contain chromosomes of two chromatids. In the process of division, special microtubules ( spindle fibers) composing the spindle apparatus are attached to the chinetochore region of the chromosomes. Motor proteins moving along these microtubules separate the sister chromatids from each other, so they halve the DNA content of the nucleus. In animal cells, spindle fibers originate from the region around a pair of cylindrical organelles, the centrioles (two centrioles compose the centrosome). From the majority of plant cells this organelle is missing.
The plane of the common wall between the future daughter cells is determined in a rather early stage of division (see below). Should this plane be in the equator of the cell, two identical cells are produced. This is equal cell division. Otherwise the daughter cells are of different size – unequal cell division. The development of several specialized cells and simple tissues (e.g. tirchomes, stomata) is initiated with an unequal division. The first division of the zygote is also unequal (for details see Chapter 4). A further characteristic of cell division is the angle of the newly formed cell wall to the nearest surface of the organ. If this wall is in right angle to the surface (i.e. it is perpendicular to it) the division is called anticlinal, while if the cells divide parallel to the surface, the division is periclinal.
1.8.4.1. Mitosis
In the process of mitosis, the DNA content of the mother cell is divided between the daughter cells without the reduction of the chromosome number, thus the ploidy of the cells does not change. (Mitosis of a haploid cell results two haploid cells, that of a diploid cell produces two diploid ones etc.) For this purpose, the sister chromatids of the chromosomes are separated from each other and later they get into different daughter cells ( Figure 11). Since the nucleotide sequence of the sister chromatids is completely identical, the genetic information within the produced cells is also the same, and it is also identical with that of the mother cell. (Consequently, no genetic recombination occurs during the process.)
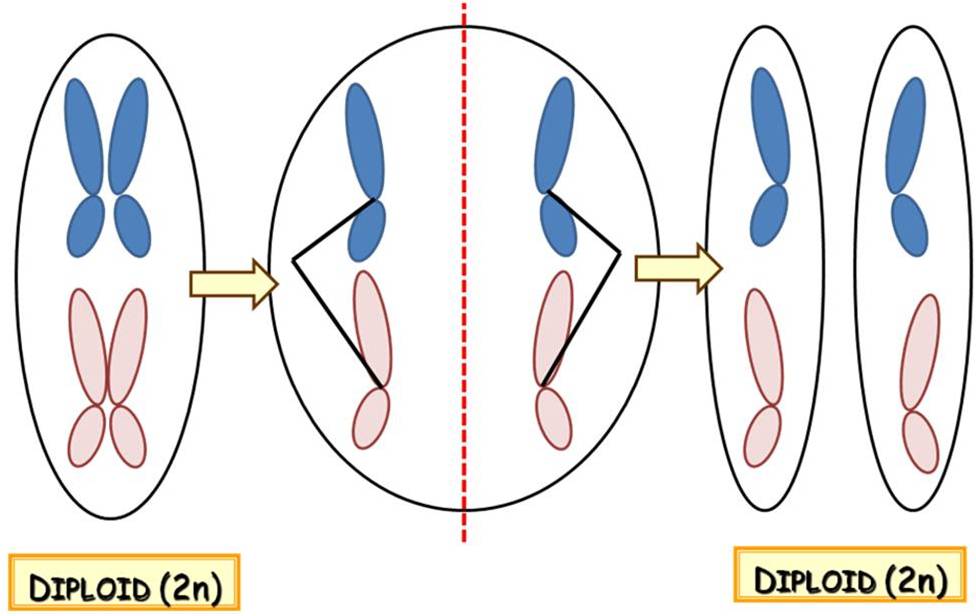
Figure/Animation 11. Overview of mitosis.
Vegetative cells of plants are produced via mitosis (within both the gametohyte and the sporophyte stages of the life cycle), just like the gametes are.
1.8.4.2. Meiosis
During meiosis, the chromosome number of the cell is halved, thus the homologous chromosomes get into different daughter cells, not only the sister chromatids are separated. This type of division has two main stages (Figure 12). In meiosis I homologous chromosomes are separated, thus a diploid cell containing chromosomes of two chromatids produces two haploid ones still bearing chromosomes with sister chromatids. Meiosis II results four haploid cells with chromosomes of one chromatid, so in this stage sister chromatids are separated from each other.
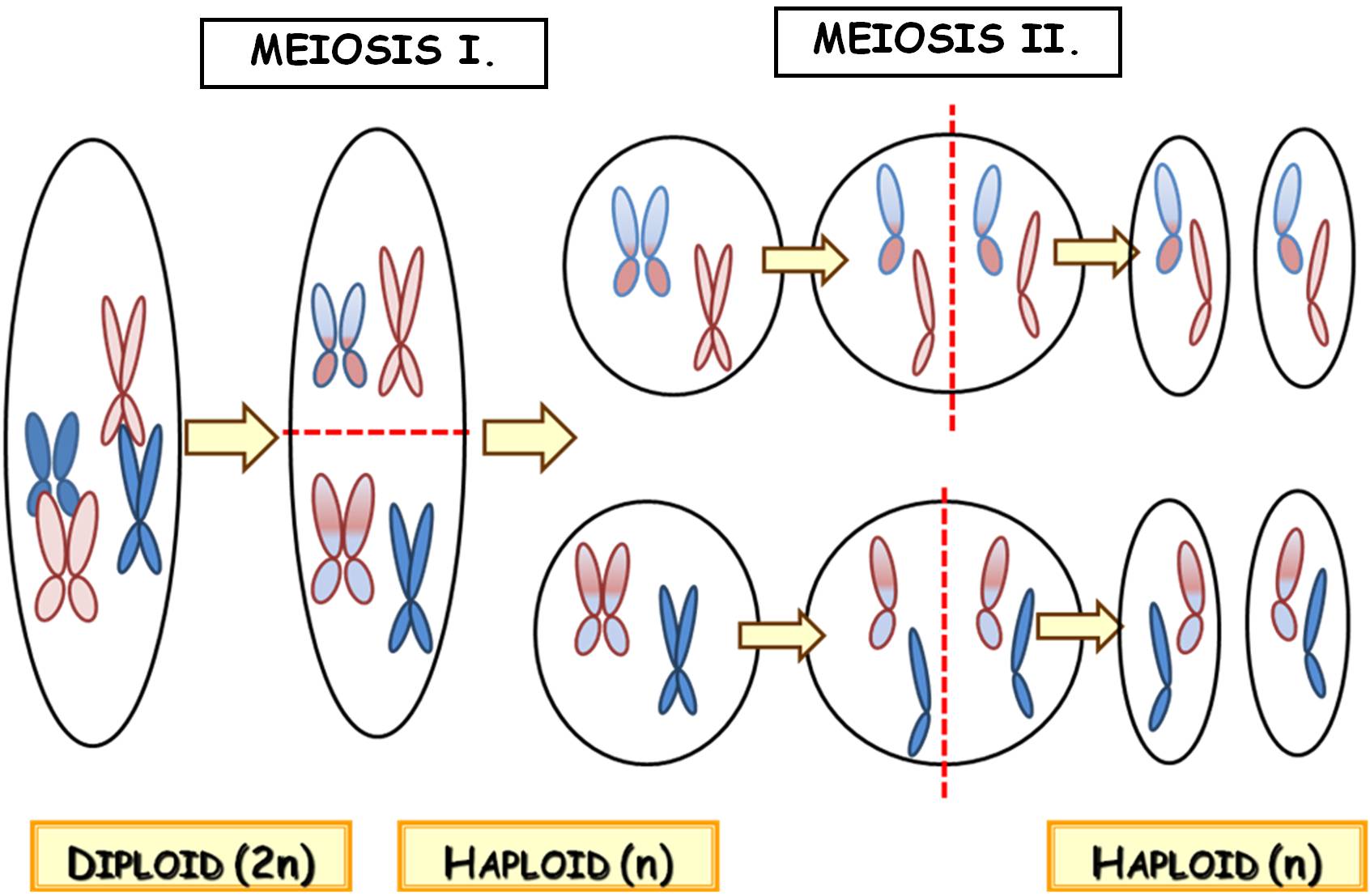
Figure 12. Overview of meiosis
Since by the end of meiosis the members of the homologues get into different daughter cells, the genetic information of these haploid cells is inevitably different. Homologues of different origin (i.e. from the egg or the sperm cell) are randomly sorted between the daughter cells in meiosis I, thus they contain a new and random combination of maternal and paternal alleles, so they are recombinant cells. The main purpose of meiosis is to produce such recombinant cells, and to maintain genetic diversity within the species.
Recombination happens not only concerning the alleles of different chromosomes, yet the maternal and paternal versions of alleles can be shuffled between the homologous chromosomes, as well. This process is the crossing over (crossover), when the chromatids of the homologues pair up in the first stage of meiosis I and the alleles of the same locus are exchanged via regulated double-stranded breaks of the DNA. Consequently, after crossing over a single chromatid of the chromosome contains both maternal and paternal segments (i.e. alleles).
In plants, meiosis produces the first cells of the haploid stage of the life cycle, i.e. the spores. It is worth to emphasise once more, that contrary to animals, gametes of plants are the final cells of the gametophyte, and so they are produced by the division of haploid cells, consequently by mitosis!
Table 3. Comparing mitosis and meiosis.
|
Mitosis |
Meiosis |
|
Ploidiy of the cell constant |
Chromosome number halved |
|
Genetic information of the daughter cells is identical and also the same as that in the mother cell (no recombination) |
Genetic information of daughter cells differ, and also differs from that of the mother cell |
|
2 daughter cells produced |
4 daughter cells produced |
|
DNA halved in one stage: separation of the sister chromatids |
DNA divided in two stages: meiosis I.: separation of the homologues; meiosis II: separation of the sister chromatids |
|
Vegetative cells, gametes |
Spores are produced via mitosis |
Table 4. Overview of the most important organelles of the eukaryotic cell .
|
Organelle type |
Organelle |
Main functions |
|
Ground substance of the cell |
cytoplasm |
medium for metabolic processes (e.g. fermentation, glycolysis, protein synthesis on ribosomes), inner transport processes and ordering of organelles (cytoskeleton) |
|
Organelles of bordering |
cell wall |
defence (against mechanical, chemical impacts, turgor and pathogens), determining volume and shape |
|
cell membrane |
integrity of the cell, signal perception, transport, |
|
|
Membrane organelles |
endoplasmatic reticulum |
lipid synthesis, detoxification (SER), synthesis of secreted proteins (RER) |
|
Golgi apparatus |
synthesis of polysaccharides (pectins, hemicellulose), modification, sorting and packaging of proteins for secretion |
|
|
vesicles |
transport between membrane organelles within the cell |
|
|
vacuole |
protein degradation, fluid accumulation (water, organic acids), sequestration of compounds disturbing the metabolism of the cytoplasm, accumulation of crystals |
|
|
Energy producers |
plastid |
photosynthesis (chloroplast), storage of carotenoids (chromoplast), accumulation of storage compounds (leucoplasts) |
|
mitochondrion |
anabolism (citrate cycle, terminal oxidation) |
|
|
Information centre |
nucleus |
storage of genetic material (DNA), site of replication, transcription and gene regulation |
References
Beck CB (2010): An Introduction to Plant Structure and Development: Plant Anatomy for the Twenty-First Century. 2. kiad. Cambridge University Press, Cambridge,
Darnell J, Lodish H, Baltimore D (szerk.) (1986): Molecular Cell Biology. Revised Printing with Expanded Index. Scientific American Books, New York.
Evert RF, Eichhorn SE (szerk.) (2006): Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body – Their Structure, Function, and Development, 3. kiad., Wiley-Interscience, New Jersey.
Gyurján I (1996): Növényszervezettan. Egyetemi jegyzet. ELTE Eötvös K., Bp.
Haraszty Á (1998): Növényszervezettan és növényélettan. 6. kiad., Nemzeti Tankvk., Bp.
Jacob F, Jäger EI, Ohmann E (1985): Botanikai kompendium. Natura, Bp.
Kovács János (1999): Sejttan. – In: Sass Miklós, Zbory Géze (szerk.): Összehasonlító anatómiai előadások I. ELTE Eötvös Kiadó, Bp.
Láng F (szerk.) (1998): Növényélettan. A növényi anyagcsere. ELTE Eötvös K., Bp.
Mauseth JD (2008): Plant Anatomy. The Blackburn Press, New Jersey.
Sárkány S, Szalai I (1966): Növénytani praktikum I. Növényszervezettani gyakorlatok. 3. kiad., Tankvk., Bp.
1 It is worth to note that although fungi cannot produce organic matter from inorganic carbon compounds (e.g. from CO2), they can build inorganic nitrogen forms (e.g. NH4+, NO2-, NO3-) into nitrogen containing macromolecules.
2 Cytological literature distinguishes the term ‘cytoplasm’ from ‘cytosol’. In this interpretation, ‘cytosol’ means the total protoplasm of the cell composing of the watery ground substance, the ‘cytosol’, together with the organelles within. (That is: cytoplasm = cytosol + organelles.)
3 In addition to secreted proteins, proteins of the lysosomes and those embedded in different membranes (integral proteins) are also synthesised here.
4 As a matter of fact, plant body is divided into well defined ’symplastic domains’ during the ontogeny. The main purpose of this phenomenon is the possibility of independent regulation of the certain domains.
5 The majority of chlorplasts contain spherical lipid bodies, plastoglobules. Previously these were considered to be mere lipid storage particles, yet recently they are proven to contain also the enzymes of certain metabolic reactions (e.g. those of the tocopherol synthesis).
6 It should be noted that another concept of chromosome is applied in genetics. According to this, chromosome means the entirety of genes comprising a common linkage group. From this point of view, we can also talk about circular bacterial chromosomes, though no histones are attached to the prokaryotic DNA.
7 When discussing the examples of this chapter, we simplify the explanation by proposing that the mentioned traits are determined by the alleles of single genes. Actually, several feature (e.g. flower colour) has polygenetic heritance, i.e. the observed trait is determined by the interaction of the alleles of more than one gene.
8 The heredity of the nuclear genome fundamentally differs from that of the organellar genome. While information coded in the nucleus derives equally from the two parents, the plastids and mitochondria of the zygote derive only from either the egg or the sperm cell (so the information coded in the extranuclear genome is determined only by the genetic material of one parent).
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028