3. Growth regulation of ornamental plants
Authors: István Dániel Mosonyi – Márk Steiner
It takes for granted to pass by a flower shop, that we can see plants in flowering in its display window all of the year, however it is well known, that the ancestries of this culture species bloomed only in a definite period of the year. The planning and the programming of the cultivation are really momentous in the ornamental plant growing the same as the other part of plant growing: so becomes possible to produce plants in the adequate size, form and advanced state for the adequate time. The growing and the developmental direction of plant organs can be affected by two different ways, but thereunto it has to be known the environmental demand, the live circle and courses of the plant. Either of this two opportunities is the indirect method. In this case the development of the plant is influenced by the changing of environmental factors, and it takes effect on biochemical processes in the plant organ. The other possibility is the direct method, when it is encroached straight into the physiological processes by chemical treatments. Both methods are wide used in the modern ornamental plant production.
3.1. Growing stages of ornamental plants
The mastery of growing stages is crucial for the adequate application of growth regulation. The development of the higher plants is divisible for two separate periods: for the juvenile (or vegetative) and for the adult (or generative) stage. The main difference between the two stages that the plants are not able to bloom in juvenile phase. It may be further peculiarity of the juvenile stage the differing leaf-form (heterophylly), the differing growing character (inclination for tillering and sprouting, or raising of creeping stems on the ground), and the more effective protection against the harmful animals (thorns, prickles, stinging-marginal leaves, foliage high in repellent fragrance). The tissue-conformation of the plants is also differed in the juvenile stage compare to the latter stadium. The stem of a juvenile plant has as usual flimsier and more parenchymatic tissue. Therefore the adventitious roots can initiate easier, so the vegetative propagation becomes faster. Contrarily while the adult phase the plants attain their whole advance state, and they are already able to grow flowers and fruits, if the environmental conditions are suitable for there. The other characteristics of the adult stage may be the changed leaf-form and growing character (non-tillering, non-sprouting, soft-growing), the loosing of the protection organs, and in the stem the stereome dominates against the parenchymatic tissues, so the regeneration ability of the plant decreases.
The two phases follow each other every time, but the transition can drag for years, especially in woody plants. This procedure takes only few weeks or months in the herbaceous plants and short-life species. The transition is affected by more factors; the most important is the plant-size. It can be specified also in absolute unit (e.g. in the case of bulbous plants, where the adult age can be defined by the diameter of the bulbs), but in the most cases the absolute volumetric sizes are not object, but the number of internodes and leaves. It can be so possible, that the plant, which was treated by internode-extension inhibitor (growth retardant), is able to bloom. The conformation of growing stages is also influenced by the water- and nutrient-supply of the plant. The vegetative organs of a well or too much supplied plant growths significantly, so the juvenile phase elongates. First of all nitrogen generates this effect, while the dominant potassium and phosphorus push the plant development to generative direction. The stinting of nutrient and irrigation induces the early ageing of plant organism, and it also means that the plant comes faster into production. This phenomenon called neoteny is a defensive mechanism in the life of the plant, because it tries to ensure by any means the advanced reproduction.


Juvenile and adult leafs of Hedera helix

Juvenile and adult leafs of Morus alba



Juvenile, transitional and adult leafs of Monstera deliciosa
But the reduced supply worsens the quality of flowering, so it has to be carefully with the using of this blooming induction method. It has to find the point where the decreased supply causes only a minimal blooming quality-loss. Therefore, the plant gets abundant supply again after the induction of generative phase, because after a successful induction the plant does not return easily to vegetative stage. However, there are some cases when the backspace to the juvenile phase can be stimulated, it called rejuvenilization. Of the vegetative propagation the cuttings and what is more the grafting have also rejuvenilization effect. The hormonal balance changes in the cuttings after the isolation from the mother-plant, and they come back again more or less into juvenile stage. In the case of grafting due to the juvenile rootstock the scion becomes also juvenile again. But this phenomenon may depend strongly on species and varieties. Sometimes the plant does not return to juvenile phase after vegetative propagation. This knowledge is exploited by the horticulturists, e.g. in the case of ’Arborescens’ type ivies where the aim is the preservation of the adult attributes on the propagated plant, too. The stronger and most safety rejuvenilization effect can be realized by external given hormones (cytokinins), which are commonly used also in micropropagation. There can be observed on woody plants, that it is on the same specimen juvenile and adult parts, too. It is due to the inequality of the light distribution. The marginal part of the crown, which has a lot of light, becomes faster adult compare to the overshadowed, lower part of the crown. This shoots in the shadow retain their juvenile form for a longer time, which advantage is taken of by the horticulturists, and they collect the cuttings from this part of the plant. So, the conformation of growing stages is a complex system in the plants which depends on more factors. The changing to adult phase does not mean automatically the starting of the flower-induction. It eventuates, if the adequate environmental conditions are realized, which will discussed at large in the next subsection.
3.2. Growing and flower-induction influencing environmental conditions
In the tropics the plants are in growth continually, because the temperature and the water-supply can be considered constant. So, the tropical plants have no dormancy period, their vegetative growing is persistent, the foliage change constantly (not simultaneously), and there are as usual flowers and fruit as well on the plants at the same time. They are the permanently-growing species, our potted foliage plants, like fig trees, philodendrons and Rex Begonias. The life-cycle of the plants on the other clime of the Earth is determined by the yearly changing periods of the seasons. This life-cycle follows closely the different weather periods, so the plant can gear its vegetative and generative life-activities to the favourable period, while the unfavourable season it can pull through reservedly. They are the fractionally-growing species, like the deciduous woody plants in the temperate zone: their buds push out when the temperature raises in the spring, the shoots grow to the middle of summer, and then they stop it. The ripening of shoots and the developing of buds occur till the autumn, then the foliage fall down, and the plants pass the freezing wintertime in leafless-status. The spring bulbous plants have two dormancies during a year: due to the drought in summer and the cold in winter. The root cause of the dormancy in the subtropical zone is not the cold, but the drought season. Beside the temperature and the water-supply the light is the other important environmental factor, which plays a role in the regulation of the plant-physiology.
Temperature
It is well known, that every plant-species have a temperature range, where the physiological processes can take place in order. This range can be described by a minimum, optimum and maximum value. It can differ from the optimum value usually with ± 4-5 °C without significant changing in the production. If the temperature is too low, the growing slows down, than it stops. It speeds up, if the temperature is too height, but the respiration-rate increases beside the constituent processes, till by and by the plant will apply more energy, than it can produce, and it will finally weaken. The temperature requirement of the permanently-growing species is 25 °C or higher. Contrarily, the fractionally-growing species have two kinds of temperature-demand: one in the growing season and one other during the dormancy.
The temperature requirement during the growing season is determined by the origin of a plant. In the case of the temperate zone species it is about 16-20 °C, while the plants stem from warmer zones it can be 25 °C or higher like the demand of tropic zone plants. The biological zero degree shows that minimum temperature, at where the plant spouts out. It is around 10 °C in the case of a lot temperate zone plants. The active temperature value is the difference of biological zero degree and effective mean temperature. The plants detect usually the accumulated temperature, namely the sum of active temperature value during a period, and they can step across to another growing stage, if this value hits an adequate level.
The temperature-demand of dormancy hangs on its type (winter or summer dormancy) and on the origin of the genus. The cold period is necessary to the plants, which needs have winter dormancy for the restarting of the growing or for the flower-induction. It is called vernalization period. Not only its temperature is important, but the time of the period is also dominant for the plants. The optimal temperature of summer dormancy equals as usual with the mean temperature of the growing season, it is about 20-25 °C. It can be also instrumental in effect of flower-induction, which opportunity is made use mainly in the case of bulbous plants. The true dormancy is often followed by secondary dormancy. It occurs, if the plant obtains the physiological state that it is ready to sprout out, but the external conditions, like the too low temperature or the drought, leave the plant in rest. It will able to exit in reality from the rest only then, if the environmental conditions are adequate for it. This is very good opportunity for the growers to time the blooming of some bulbous plants.
The circadian rhythm of temperature (changing of daytime and night time temperature) has also important role in the growing of plants. In the growing used, so called DIF parameter means the difference between the daytime and night time temperature. Stem elongation is promoted by warmer days than nights (positive DIF) and inhibited by warmer nights than days (negative DIF). Plants grow taller when DIF becomes more positive and plants remain short as DIF becomes smaller or more negative. But not every plants show the same reaction for DIF’s changing. The plants originate from temperate zone respond stronger, than the tropical species. This is because of the difference between the two climate conditions.
Light
Three responses occur in the plant due to the light-exposure: the photosynthesis, the photomorphogenesis and the phototropism. The photosynthesis, like constituent and assimilation metabolism is not detailed here, but the photomorphogenesis (light-induced ontogeny processes) and in smaller compass the phototropism (motion controlled by light stimulus) have an important role in the growth-regulation. The plants can realize three attributes of the light: its wavelength, intensity and the photoperiodism (the length of light stimulus). The wavelength range utilized by the plants coincides more or less with the visible light range, but inside this range the bioavailability of the difference colours is not equable. The chlorophyll molecule is able to use the red range during the photosynthesis (in a small compass the blue range – between 440-470 nm – is also useful), while the flavin and carotenoid pigments use the blue range, too. The yellow and green ranges have less notability, but they also have some physiological effects. The red light has also the undermentioned physiological effects: stimulates the germination, the stem expansion and the dry matter accumulation. It enhances the synthesis of anthocyanins, and helps to step over to the generative life stage. The red light has effect also on hormonal regulation, because it is essential for the induction of organogenesis, because it helps to accumulate the cytokinins in the tissues, and so induces more branching. At the same time it reduces the volume of auxins. The infrared radiation (above 735 nm) results really stong stem elongation on the plants. Contrarily, the blue light reduces the long of internodes, and so makes stronger the stem, and enhances its resistance, as well as it makes better the colour of the flowers and the leaves. Due to the blue light increases the biosynthesis of gibberellins and abscisic acid. The green and yellow ranges have smaller effect, but it is demonstrable, that the degradation of auxins is significant in the case of illumination in this ranges. Therefore, relative cytokinin dominance realizes in the tissues, and it has favourable effect on secondary shoot induction and on development of buds.
The march of the seasons is shown for the plants by changing of light intensity and the daily length of lighting. So, the life-cycle of plants can adapt to the periodical alteration of environmental conditions. The answer of the plants for the changing in the ratio of day and night is called photoperiodic reaction. It has notability particularly in the flowering induction. The photoperiodic reaction-time is a typical of genus and variety duration. During this time the plant gets to the whole blooming from the starting of the photoperiodic stimulus. Its length is been in terms of weeks. Sometimes, the length of reaction-time is modified by the changing of light intensity. Thus, it can be talked occasionally about accumulated light, similar to accumulated temperature. However, it is not true universally, because in the case of the most plants the shortening of the duration period is not supplemented by the increased light intensity. The specimens of a plant genus start to bloom over or below the critical day length. The values below the critical day length are called short-day in the literature, while values of the long-day are over the critical day length. (These values are between 10-15 hours.) Short-day ornamental plants flower when the day lengths are less than their critical photoperiod. Otherwise, there are long-day plants. The short-day plants bloom in the nature by autumn, while the long-day plants during the summer term. Actually, not the day length, but the night length is important for the plants. It is confirmed by the fact, that in the case of short-day plants the blooming evocative effect of long-night can be broken by a minimal use of artificial light. A few minutes are enough to knock out the effect of long-night. The plants are reactive in both groups to light stimulus obligate (qualitatively) and facultative (quantitatively). Those species, which have obligate reaction, start to bloom alone then, if they get the adequate stimulus from the length of dark period. Contrarily, the facultative reaction plants start to bloom sooner or later also without light stimulus, but the flowering will be slower compare to the specimens, which grow on adequate day length conditions. Furthermore, it is important to know, that there are also day-neutral plants, where the blooming date is completely regardless of the changing day length.
A Ornamental plants with phothoperiodic reaction
|
obligate long-day |
facultative lomg-day |
facultative short-day |
obligate short-day |
indifferent |
|
|
Cut flowers, pot plants |
cut carnation, Eustoma grandiflorum |
Begonia elatior hybrids |
Chrysanthemum , Kalanchoë, Euphorbia pulcherrima |
Gerbera , Rosa, Strelitzia, Freesia, Pelargonium, Saintpaulia, Primula |
|
|
Annuals |
Centaurea , Linum, Gazania, Lavatera,, Lobelia, Mimulus, Oenothera, Begonia × tuberhybrida, Fuchsia hybrids, Rudbeckia |
Ocimum , Viola, Salpiglossis, Ageratum, Calendula, Dianthus chinensis, Tithonia , Petunia Grandiflora típus, Salvia farinacea |
Celosia , Sanvitalia, Zinnia elegans, Cosmos, Gomphrena, Ipomoea, Tagetes tenuifolia |
Tagetes erecta , Lablab purpureus, Ipomoea lobata |
Amaranthus , Cleome, Tagetes patula, Impatiens New-Guinea hybrids, Impatiens balsamina, Begonia semperflorens, Zinnia angustifolia |
3.3. Taking part chemicals in the plants during growing and flower induction
The communication between cells takes place by the help of chemical messengers, same as in an animal organism. The plant hormones are a specific type of these chemicals, in a wide sense they are growth regulators. They take effect by the connection to special proteins, to the receptors. The hormones influence first of all on growing and development. Their concentration is very low, but their action depends on the volume of concentration, and their effect evolves far from their origin place (expect the ethylene, which can arise in every cells, and it effects on the spot). One phenomenon is controlled by more hormones, so beside the concentration, the interaction of the different hormones has also significant effect on final results. The hormones have inhibitory and stimulating effect, too. Traditionally five types of hormone can be separated: the auxins, the gibberellins, the cytokinins, the ethylene and the abscisic acid.
Auxins
The indole-3-acetic acid (IAA, Hungarian abbreviation: IES) fills the part of auxin generally in higher plants in vivo. Other indole compounds (indole-3-butyric acid or IBA, Hungarian abbreviation: IVS, 4-chloro-IAA, OH-IAA) have similar effects in the plant like IAA, but they are usually the precursors or derivatives of IAA, and they become IAA in the plant organ. There are also synthetic auxins, which have auxin-like effects, but they do not metabolise to IAA. Because the decomposition products of these compounds have also auxin-like effects, furthermore they are more resistant against enzymatic dissociation, their influence keeps in the plants for longer time. Accordingly, the horticultural use of them gives more effective results, like IAA, which is anyway a light sensitive compound, therefore it is difficult to store and use it.
The most important synthetic auxins:
- 2,4-dichlorophenoxyacetic acid (2,4-D),
- 2,4,5- trichlorophenoxyacetic acid (2,4,5-T),
- 1-naphthylacetic acid (NAA),
- 2-methyl-4- chlorophenoxyacetic acid (MCPA),
- 4-amino-3,5,6-trichloro-2-pyridinecarboxylic acid (picloram),
- 3,6-dichloro-2-methoxybenzoic acid (dicamba).
The physiological effects of auxin can be summerised as follows:
- it stimulates the extension growth of cells and organs especially of the stem,
- it advances the growth of lateral roots and the root induction of shoot and leaf cuttings,
- it plays a determining part in the apical dominance,
- it regulates the exfoliation of leaves and fruits, the binding, growing and ripening of fruits,
- it interacts with other hormones, too: it stimulates the cell division together with cytokinin,
- together with gibberellins it can influeces the sexual character of flowers (the high auxin/gibberellin ratio results more female, otherwise more male flowers).
The auxin is produced in the plants by the leaves and growing tip, and it can be translocated downwards in the phloem.
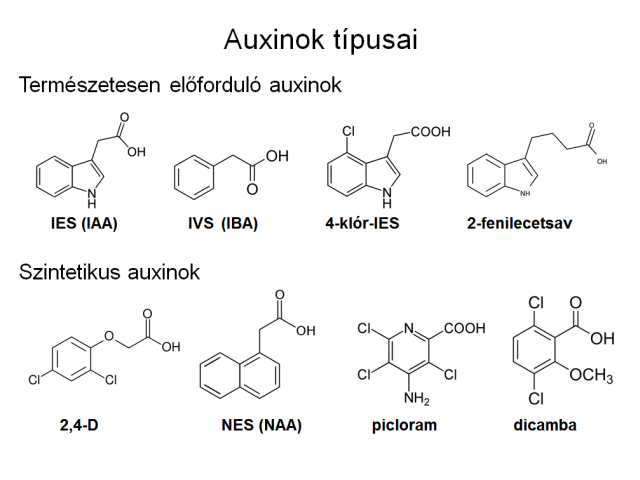
Types of auxins (natural and synthetic)
Gibberellins
Gibberellins were found not only in higher plants, but also in lower plants, bacteria, algae, and fungi. Gibberellins are tetracyclic diterpene acids, currently more than 100 kinds of gibberellins are known. They are synthesised in the stem tip, and they can be translocated in xylem and also in phloem and they occur in every plant organ.
Their effects somewhat overlap with the auxin:
- stimulation of stem extension,
- induction of parthenocarpy,
- amplification of apical dominance.
But they have specific effects, too, which can not be generated by auxin. These are the followings:
- normalisation of genetic and physiological dwarfism,
- germination induction of light- and cold-demanding seeds,
- breaking of bud dormancy,
- promotion of blooming, or inchoation of them in non-inductive environment in the case of plants which need vernalization or long-days,
- alteration of the sexual character of flowers towards the male character.
The most commonly used gibberellin in the horticultural is the GA3. It is applied by spraying for hitting the following target: induction of parthenocarpy on grapes, yield increases on seedless varieties, which have low productivity, induction of blooming in non-inductive conditions. The soaking method is used for the facilitation of germination. The gibberllin induced flowering is possible only on those long-day plants, which have rosette under short-day conditions, and the stem extension and flower formation eventuate under long-day conditions. So, on these plants the gibberellin can substitute the long-day treatment. On those plants, which do not form rosette (e.g. the fuchsia), the gibberellin is not able to promote the blooming. Furthermore, the gibberellin treatment has effect on short-day plants neither in positive nor in negative direction. The gibberellin treatment can be adapted for the substitute of vernalization, but this phenomenon is still not confirmed generally, because there are so cold-consuming species, which did not get flower after gibberellin treatment. In the case of bulbous iris the GA3 treatment substitutes the vernalization right, if the plants get the adequate light period after treating (it is necessary also then, if there is no GA3 treatment, only vernalization).
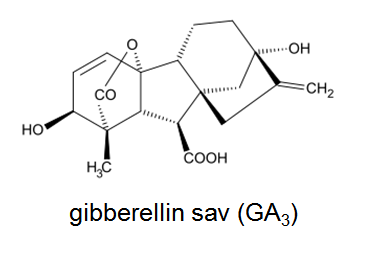
Structure of gibberelic acid
Cytokinins
Similar to gibberellins the cytokinins are also present both in higher plants and in fungi and bacteria. It shows their general incidence, that the bacteria have the same cytokinins compare to higher plants, or rather they are analogue of them. The collective noun means purine structure compounds, which induce cell division, and they maintain the persistent growing of tobacco callus culture in the attendance of auxin. Different cytokinins are found naturally in the plants, but several synthetic types are also known. The zeatin, isopentenyl-adenine, and 6- benzylaminopurine (BAP), and its hydroxylated forms (ortho- and meta-topolin), which are typical in Populus species, are found naturally in higher plants. The 6- benzylaminopurine (BAP) (or 6-benzyladenine, BA) is registered also as synthetic cytokinin, same as to kinetin. The cytokinins are produced in the plants by the root apex, and they are transported in the xylem, and they get to the aboveground part of plant with the transpiration flow.
Their effects include a lot of physiological processes:
- they participate in the regulation of cell division, in growing, developing and differentiation processes,
- they stimulate the cell extension,
- they start the germination without light effect in the case of seeds, which need red light for germination,
- in the case of some species, which need vernalization for blooming, cytokinin treatment can substitute it,
- they delay the leaf senescence,
- they stimulate the nutrient mobilisation,
- they act as the antagonist of auxin in the course of mechanism of apical dominance,
- they stimulate the bud growing,
- they advance the tuber formation,
- they stimulate the chloroplast maturation,
- they induce the enzyme synthesis.
The cytokinins are utilized in the horticultural beyond the micropropagation for the promotion of branching in the case of woody plants (products containing benzyladenine). Furthermore, the cytokinins dissolved in water slack the droop of flowers, the leaves remain greener, and the vase life get well in the curse of postharvest preparation of cut flowers.
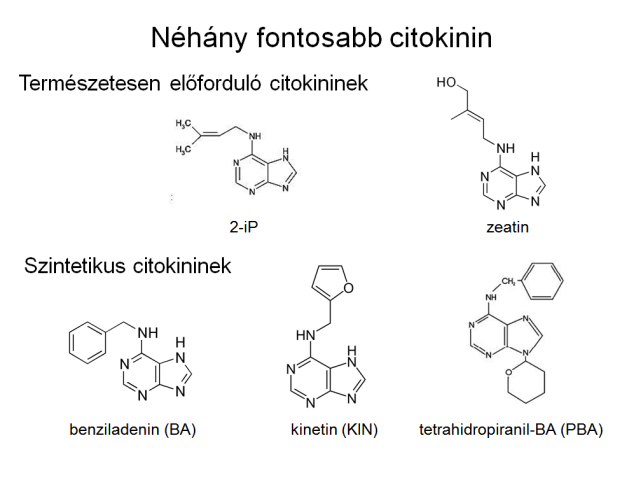
Structure of some cytokinins
Abscisic acid
In the 1960s a growth regulator, which proved to be the same in every case, was identified from more plants, and forasmuch it was supposed, that this compound causes the abscission of the plant organs, it was named after for abscisic acid (ABA). Today it is known, that the ethylene is accountable mainly for it, but the denomination remained behind. In higher plants this compound is detectable in every growing phase, but only in very small quantity. Contrary to the previous hormone groups this is an inhibitor, a regulator compound with inhibitory effect, however its notability is the same compare to them. The physiological effects of ABA:
- it takes part in the leaf abscission, and in the conformation of abscission zone,
- it develops or rather abolishes the bud dormancy together with other growth regulators,
- it develops or rather abolishes the seed dormancy, the external application has anti-germination effect,
- it accelerates the senescence,
- it enhances the adventitious root formation,
- it inhibits the extensional growing,
- it induces blooming on some short-day plants in non-inductive circumstances (strawberry, blackcurrant),
- in the case of water deficit its concentration increases, it enhances the water uptake of roots and deflates the transpiration due to the closing of stomata.
ABA could be widely used in the horticultural practice, but since it is metabolised quickly by the plants, it should be applied more times and in higher concentration, which is not profitable due to its high prime cost.
Ethylene
Ethylene is also called as hormone of senescence and stress. Every cells of a plant organism can produce it, mainly under abiotic stressors: cold, drought, injury, absence of air by the roots, high ozone and heavy metal concentration. It is produced in a higher volume by the senescent organs and fruits, than by the mature, vegetative tissues. Its physiological effects are the follows:
- it inhibits the extensional growing,
- it enhances the gibberellin sensitivity of shoot,
- it stimulates the leaf and fruit abscission,
- it induces rooting,
- it breaks the bud and seed dormancy,
- it stimulates the flower initiation, the fruit setting and growing,
- it alters the sexual character of flowers towards the female character,
- it advances the tuber formation,
- it accelerates the senescence,
The horticultural use of ethylene is significant. It is a gas, so it could easily evaporate, therefore it is applied not in direct form, but such compounds are used, from which the ethylene disengages after decomposition (e.g. active substance 6-chloroethylphosphonic acid in Ethrel and CEPA). Forasmuch, it accelerates the overblowing of cut flowers, therefore the concentration of the permanently producing ethylene has to be declined in the storage room by its absorption. To this end, a filter made of porous, large veneer material is used, which is saturated by potassium permanganate solution. At the same time, ethylene is important implement for early flower induction by bromeliads.
3.4. Other growth retardant, dwarfing compounds
A special group of growth retardant, the so-called dwarfing compounds is widely used in ornamental plant growing. In the conventional sense they are not plant hormones, but synthetic compounds. The market demands the little sized plants with compact habit, which production can be solved in absence of adequate genotype often only by chemical application. Those compounds are employable for this purpose, which are the antagonist of gibberellin, which generates stem/internode elongation. They inhibit the gibberellin biosynthesis, or they balk the apical dominance and result in bushy plant habit. Furthermore, this compounds increase the number of flower buds, but they can delay the blooming by 1-2 weeks. The following triazole-type antifungal agents are adapted for this goal: paclobutrazol, ancymidol, flurprimidol, daminozide, and chlorocholine chloride (CCC). However, due to the increased environmental regulation the number of available compounds, which have these agents, declines more and more.
3.5. Opportunities of practical execution
Optimisation of propagating time
This method can be used for blooming regulation, if the plant blows only after an adequate time or plant size. In the case of pretreated (heat treated) propagation material the Amaryllis is a good example of it. But in the most crops the optimisation of propagating time is combined with other growth regulation opportunities.
Mechanical treatments
The mechanical treatment is on the one hand a physical stimulation of plants, on the other hand removal of some parts of plant. The physical stimulation means the sweeping of the growing crop, which is solved by a plastic or wooden slat, perchance by brushes over the crop. The shoot tips of plants are swept over several (30-40) times per a day. Its effect is similar to the inflection of wind: the longitudinal growing slows. It can be used alone on healthy plants, taking care not to do any injury. This method is not current in Hungary, but its permeation is probable, because its need for investment is low and it needs no compounds, which have harmful effects on environment. The changing of shoot position (the bending method, which is used often for cutting rose production in greenhouse) has stronger physiological effect compare to physical stimulation. The large changing of shoot position is made by the refracting, twisting and bending down of the growing shoots above their few lower buds. This effects the termination of apical dominance in the plant, and the dormant buds in the leaf axils at the base of the shoot (below the bend) begin to grow. The abolition of apical dominance is also resulted by pinching, cutting back and breaking out in different plant height. The treatment will be properly effective, if not only the shoot tip will be removed from the plant, but also the one or two youngest leaves. In this case the outgrowing will be adequately fast from the axillary buds, and better tillering will be realized, compare to the simple shoot tip removal. Cutting back and breaking out up to older part of plant are applied more rarely. They are used, if it is late for the adequate time of pinching. In such case the outgrowing of axillary buds is more languid, and less of them will grow out.

Developing of axillary buds of Poinsettia after pinching
The attributes of a pot (especially its capacity) determine the maximized root mass. This root mass can supply only a limited above-ground plant size with nutrient and water, so the potential size of plant will predestinate. Choosing the size of the pot it is necessary to be aware of, that if the plant reaches the pick of its size, it can easily senescence.
Short-day treatment
It is made by blackout, and it can be needful in Hungary from April to September. The growing crop is blanketed by opaque material. Usually densely spun stuff is applied, which is stretched on a frame-system over the plants. The black plastic cover is also applicable, but it has to be careful, because the plants are not able to ventilate below the foil, and the crop can heat up extremely. It can be got around easily by a special foil, which outside has an aluminium film. It reflects the heat, and so encumbers the over heating. If the frame-system has sufficient quality, the covering material can be used as energy curtain to pare down the expenses of heating season.

System for short-day treatment in production of Kalanchoë blossfeldiana

System for short-day treatment in production of Poinsettia
Supplementary assimilation lighting
It is applied for the reaching of higher yield in the case of light-demanding and facultative long-day plants (e.g. rose, lily). Its minimal intensity is suggested around 4000-5000 lux in the most ornamental crops. The lower light intensity under a long period is more efficient, than a sort but intensive lighting (e.g. a 24 hour long lighting on 4000 lux is more effective compare to a 12 hour long supplementary lighting on 8000 lux). It is used in Hungary from the end of October to the beginning of March. The daytime is elongated by it on a sunshiny day with 4-6 hours, while on a cloudy day the supplementary lighting can run all day long. Using 60-100 W light bulbs over the plants is enough to give long-day stimulus, but for supplementary assimilation lighting the higher light intensity and better spectrum for the plants (which are given by HPS – high pressure sodium – lamps) are indispensable. The 400 W light bulbs are the most prevalent type of HPS lamps, because their temperature is not so high yet, that it could damage the plants, if they are fixed up 3-4 m over the growing crop. This 400 W light bulbs, which emits as usual 50000 lm, are placed per 16 m2 on 4 m high (namely, the source of light projects on a 16 m2 surface from 4 m high), and so they give 50000/16=3125 lux lighting in the level of the plants.

High pressure sodium – lamps
In the future the LED light systems, which are already well-known in micropropagation, will be applied probably also in greenhouses. The great advantage of these LED-systems that the emitted wavelength range can be varied ad-libitum, and so the ratio of blue/red/infrared ranges can be changed easily in the emitted light. The using of light filters is an other opportunity in the future to regulate growing by light. In this case the natural light, which enters into the greenhouse, is filtered, and the ratios of wavelength range, which are adequate for the actual developing stage, can be generated.
Heat treatments
The growing of plants can be regulated also by alteration of temperature. For example growth inhibition can be realized by the reducing of the above-mentioned DIF-value, and the volume of chemical retardant can be so declined. Beyond the DIF-regulation the 3-6°C lower matutinal temperature (compared to night), which takes for 2-3 hours long, can give similar or better result on growth moderation. It is not needful to apply DIF-treatment through the growing season, it is enough in the most active vegetative growing period. Generally speaking, the growing time increases due to the reducing of DIF-value (if DIF reducing is achieved by the decreasing of daily average temperature – so, if it is heated less day and night).
In the case of that species, which need cold period for blooming, the length of dormancy can be influenced by holding adequate temperature. This species need often shorter cold period, than it is true in the nature, where after their active dormancy they can stay at rest because of the cold weather. During the growing the cold period can be cut down to that minimal time, while the active dormancy run its course in the plant, then they can make bloom with the increasing of temperature, earlier, than in the nature. The inhibition and timing of sprouting can be controlled by extended cold period, namely the plant is held in cold to the wanted date of sprouting, however the active dormancy is already finished (e.g. the 2-degree tulips, which already have got all stages of vernalization, and then they are held on 2 °C until bedding). It is practical to time these plants in the temperate zone so, that their vernalization period coincides with the cold season (anthurium, Easter cactus). The rate of light supply can influence the length of vernalization period – the low lighting elongates, the enough lighting cuts it down. Some subtropical species (e.g. Cymbidium) need so high light intensity under vernalization, which can not be warranted (without supplementary assimilation lighting) under our overcast weather conditions in winter, so the cold period has to be given to this plants in summer by the cooling of the greenhouse. It is very expensive, but solvable. The most current resort is the greenhouse evaporative cooling pad system. On an end wall of the greenhouse water is oozed cross a material with large surface, while ventilators on the other side draw the air cross the greenhouse. So, the temperature can be decreased due to the evaporation of water. In the case of some plants it can be talked also about heat treatment. For example the summer dormancy can be imitated in Dutch iris by holding on 30 °C, and so its bulbs may be stored for a year, until the start of next culture. In hyacinth the coalesced stem of flower is initiated by a heat treatment under the summer dormancy for better aesthetical value. The bulbs are held on 23 °C for 10 days, on 30 °C for 2 weeks, then on 25.5 °C for 3 weeks.

Greenhouse evaporative cooling pad system

Ventilators on the other side of greenhouse
Application of chemicals
The chemicals are applied first of all to stimulate the rooting of cuttings and to dwarf the plant habit. It was mentioned earlier, that the auxins (particularly the synthetic ones: indole-3-butyric acid, IBA and 1-naphthylacetic acid, NAA) are used for promoting of rooting. The advantages of synthetic auxins are the more stable structure compares to natural auxins, and they are not decomposed by heat and light. Only their sodium and potassium salts can dissolve in water, so they are applied to make aqueous solution. Otherwise, their straight powder can dissolve well only in organic solvents (e.g. in ethyl alcohol). The treatment of the cuttings can be solved by using of 0.1-0.5 % alcoholic solution (the hormone powder is dissolved in 50 % alcohol). The basal part of the cuttings is plunged into the solution for 5 second in depth of 1-1.5 cm, then the cuttings are shaken off and they are let to dry for a few minutes before sticking. Instead of solution the other opportunity is the powder preparation, which is available as manufactured. It is prepared by the mixing of concentrated alcoholic auxin solution and talcum powder, and it is waited for the evaporation of alcohol. The talcum powder dries, and the hormone is distributed in it uniformly. Its utilization is similar to the solution, but the bottom of the cutting has to be mildly wet before the dipping, because it is the only way to stick enough hormone powder to the cutting.
The growth retardants can be got out by irrigation and/or spraying. The irrigation needs less technical know-how, but more substances. It can be difference between plant species, that which kind of placement has the better efficiency. To keep clear of washing down it is practical to pass over the misting within 24 hour. The applied concentration depends in every case on the type of growth retardant, it can be found in the manufacturer’s information. For the most part, the plants begin to turn yellow after the treatment, which is a natural physiological phenomenon, and it can use as an indicator: it shows the uniformity of the placement. This chlorosis can be abolished by placement of high nitrogen content foliar fertilizers, which can be started 2 or 3 days after the treatment.

Ismelia carinata plants, treated with growth regulators: Non-treated (0) Caramba (1), Regalis (2), Alar (3)
Literature
- RÜGNER, W.: Virágképződés és virágfejlődés, Mezőgazdasági Kiadó, Budapest 1977
- TURCSÁNYI, G. (szerk.): Mezőgazdasági növénytan, Mezőgazdasági Szaktudás Kiadó, Budapest 2001
- HARASZTI, E.: Zsebkönyv a növények életéről, Natura, 1977
- ERDEI, L. (szerk.): Növényélettan. Növekedés és fejlődésélettan, JATEPRESS, Szeged 2004
- PETHŐ, M.: Mezőgazdasági növények élettana, Akadémiai Kiadó, Budapest 2002
- SCHMIDT, G. (szerk): Növényházi dísznövények termesztése, Mezőgazda Kiadó, Budapest 2002
- HORTOBÁGYI, T. (szerk.), SIMON, T. (szerk.): Növényföldrajz, társulástan és ökológia, Nemzeti Tankönyvkiadó, Budapest 2000
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028