Dísznövények növekedésszabályozása
Szerzők: Mosonyi István Dániel – Steiner Márk
Természetesnek vesszük, ha egy virágbolt előtt elhaladva annak kirakatában - legyen szó az év bármely szakáról – virágzó növényeket látunk, holott jól tudjuk, hogy ezen kultúrfajok ősei az évnek csak egy bizonyos szakában virágoznak. A dísznövénytermesztésben, csakúgy, mint a növénytermesztés többi területén is, a tervezés és a termesztés programozása nagy jelentőséggel bír a végeredmény szempontjából: így válik lehetővé adott időre adott méretű, formájú, fejlettségi állapotú növények előállítása. A növény növekedését, fejlődési irányait kétféle módon lehet befolyásolni, ehhez azonban ismerni kell az adott növény környezettel szemben támasztott igényeit, és a növény életritmusát, -folyamatait. Az egyik lehetőség, hogy indirekt módon, a környezeti paraméterek változtatásával befolyásoljuk a növény fejlődését, melyek hatására a benne lezajló biokémiai folyamatok megváltoznak. Másik lehetőségünk pedig a direkt módszer, amikor közvetlenül a biokémiai folyamatokba avatkozunk be vegyszeres kezelés formájában. Mindkét módszert elterjedten alkalmazzák a modern dísznövénytermesztésben.
Dísznövények fejlődési szakaszai
A fejlődési fázisok ismerete elengedhetetlen a növekedésszabályozás megfelelő alkalmazásához. A hajtásos növények fejlődése két elkülönülő szakaszra osztható: a juvenilis (fiatalkori vagy vegetatív) és adult (idős vagy generatív) szakaszra. A két szakasz között a legfőbb különbség, hogy juvenilis állapotban a növények nem képesek virágozni. További jellemzője lehet még a fiatalkori szakasznak a heterofília (a növény levélformája eltérő fiatal és időskorban), eltérő növekedési jelleg (bokrosodásra, sarjadzásra hajlamos növekedés, vagy kúszóhajtások nevelése a talajfelszínen), hatékonyabb védekezés az állati károsítók ellen (tövisek, tüskék, szúrós szélű levelek, repellens (távoltartó) illatanyagokban gazdagabb lombozat). A fiatalkori szakaszban a növények szöveti felépítése is eltérő a későbbi állapottól, ilyenkor többnyire lazább és több parenchimatikus szövetet (dús plazmájú, sok zöld színtestet tartalmazó szövet, mely legnagyobb mennyiségben a levél mezofillumában található, feladata az asszimiláció és a gázcsere) tartalmaz a szár, melynek következtében könnyebben alakulnak ki járulékos gyökerek, vagyis könnyebb a vegetatív szaporítás. Ezzel szemben az időskori szakaszra a növények elérik teljes fejlettségi állapotukat, melyben már képesek a virág- és termésnevelésre, ha a környezeti körülmények is megfelelőek erre. Egyéb kísérőbélyegei lehetnek az adult fázisnak a megváltozott levélforma és növekedési jelleg (nem bokrosodó, nem sarjadzó, gyengébb), a védőszervek elvesztése, a szárban pedig túlsúlyba kerülnek a szilárdítószövetek a parenchimatikus szövetek kárára, így a növény regenerálódóképessége csökken.

3.1. ábra A borostyán (Hedera helix) fiatalkori levélalakja és habitusa

3.2. ábra A borostyán (Hedera helix) időskori levélalakja és habitusa

3.3. ábra A Monstera deliciosa fiatalkori levélalakja

3.4. ábra A Monstera deliciosa átmeneti levélalakja a juvenilis és adult fázis között

3.5. ábra A Monstera deliciosa időskori levélalakja

3.6. ábra A fehér eperfa (Morus alba) juvenilis és adult levélformája
A két fejlődési fázis mindig időben követi egymást, ám az átmenet elhúzódhat több évig is, ez fás növényeknél jellemző. A lágyszárúaknál, rövid életű fajok esetében hetek-hónapok alatt lezajlik a folyamat. Az átmenetet több tényező is befolyásolhatja melyek közül a legfontosabb a növény mérete. Ezt lehet abszolút mértékegységben is megadni, – gondoljunk a hagymás növényekre, melyeknél a már virágzóképes fejlettségi állapotot a hagymakörméret alapján lehet megállapítani - , de legtöbb esetben nem az abszolút térfogati méretek számítanak, hanem az ízközök vagy levelek mennyisége – így lehetséges az, hogy egy szártagmegnyúlást gátló szerrel kezelt, ún „törpésített” növény is képes virágozni. A növény víz- és tápanyagellátása is befolyásolhatja a fejlődési szakaszok alakulását. A jó vagy túlzott ellátásban részesülő növény vegetatív szerveinek fejlődése válik hangsúlyossá, így a juvenilis fázisa általában meghosszabbodik. A tápelemek közül elsősorban a nitrogén az, amely ilyen hatást előidéz, míg a kálium és foszfor túlsúlya a generatív irányba tolja a növény fejlődését. A tápelemek és az öntözés bizonyos mértékű megvonása a növény idő előtti öregedését idézi elő, és egyben gyorsabb termőre fordulást is jelent. Ez a neoténia néven ismeretes jelenség azonban egy védekező mechanizmus a növény részéről, a növény igyekszik mindenáron a fajfenntartást biztosítani. Tudni kell azonban, hogy a csökkent ellátás miatt a virágzás minősége is csökken, így ezzel a virágzásindukáló módszerrel óvatosan kell bánni, és meg kell találni azt a pontot, ahol még az ellátás csökkentése minimális minőségveszteséggel jár a virágzás szempontjából. Ezért általában a generatív szakasz indukálása után bőséges ellátásban részesítik a növényt, hogy ismét megfelelő kondícióba kerüljön. Ilyenkor ez már nem hátráltatja a virágzást, ugyanis az indukció megtörténte után a növény már nem könnyen lép vissza a vegetatív fázisba. Van olyan eset azonban, amikor a juvenilis állapotba történő visszalépést előidézhetjük, ezt hívják rejuvenilizációnak. A ivartalan szaporítás módjai közül a dugványozás, sőt még az oltás is rejuvenilizációs hatással bír. Az anyanövényről izolált növényi részek hormonális háztartása megváltozik, melynek következtében többé-kevésbé ismét fiatalkori állapotba kerülnek a levágott dugványok, oltványok esetében pedig a juvenilis alany hatására kerül az oltott rész is juvenilis állapotba. Ez azonban erősen faj ill. fajtafüggő lehet, egyes esetekben az ivartalan szaporítás hatására sem alakul vissza a növény juvenilis állapotba, és ezt használják ki a kertészek, például borostyánok esetében az ún. ’Arborescens’ típusoknál, ahol kifejezetten az a cél, hogy megőrizzék az adult fázisra jellemző bélyegeket a szaporított növényen is. A legerőteljesebb és biztosan bekövetkező rejuvenilizációs hatást azonban külsőleg adagolt hormonokkal, citokininekkel érhetjük el, ez történik a mikroszaporítás (azonos genotípusú, szelektált fajok illetve fajták vegetatív, fajtaazonos szaporítása steril laboratóriumi, kontrollált körülmények között) során is. Érdekes jelenség a fás növények esetében, hogy egyazon növényi egyeden belül előfordulhatnak juvenilis és adult részek is, ami a fényellátás egyenlőtlenségeiből adódik. A bő fényellátottságú, a korona külső részén fejlődő hajtások hamarabb váltanak generatív fázisba, mint a beárnyékolt, alsóbb részeken fejlődők. Ezek az árnyékba kényszerített hajtások több ideig megőrzik juvenilis alakjukat, melyet a kertész ki is használ, és a dugványnakvalót ezekről a könnyebben gyökeresedő részekről szedi meg. A fejlődési fázisok alakulása tehát bonyolult, több tényezőtől függő rendszerben működik a növényeknél. Az adult fázisba történő átváltás még nem jelenti automatikusan a virágképződés elindulását is, az csak megfelelő környezeti feltételek teljesülése esetén következik be, melyet a következő alfejezetben tárgyalunk részletesen.
A fejlődést és virágképződést befolyásoló környezeti tényezők
A trópusokon folyamatos vegetációban vannak a növények, mert a hőmérséklet és vízellátás állandónak tekinthető. Emiatt a trópusi növények életében nincs nyugalmi periódus, a vegetatív növekedés állandó, a lombozat nem egyszerre, hanem folyamatosan cserélődik, a növényen általában mindig van virág és termés is egyszerre. Ezek a folyamatosan vegetáló fajok, dísznövényeink közül leginkább a cserepes levéldísznövények tartoznak ide, fikuszok, filodendronok, királybegóniák. A Föld többi táján az évszakok váltakozásával kialakuló éves periódusok határozzák meg a növények életritmusát is. Ez az életritmus szorosan követi az eltérő időjárású időszakokat, így a növény a vegetatív és generatív élettevékenységeit a kedvező időszakokhoz tudja igazítani, a számára kedvezőtlen időszakokat pedig visszahúzódva vészeli át. Ezek a szakaszosan vegetáló fajok. Jó példa erre a mérsékelt égövi lombhullató fásszárú növények életritmusa: tavasszal a hőmérséklet emelkedésével a fák rügyei kihajtanak, nyár közepéig folytatódik a hajtásnövekedésük, aztán leáll. A hajtások beérése, rügyek kifejlődése őszig megtörténik, ezután pedig lehull a lombozat, a téli fagyos időszakot a növények pedig lombtalan állapotban töltik. A tavaszi hagymás virágok életében két nyugalmi periódus is van, nyáron a szárazság, télen a hideg miatt. A szubtrópusi égövben pedig a nyugalmi periódus fő kiváltója nem a hideg, hanem a száraz évszak. A hőmérséklet és nedvességellátás mellett a fény az a másik fontos környezeti tényező, ami még szerepet játszik a növényi életfolyamatok szabályozásában.
Hőmérséklet
Jól ismert, hogy minden növényfaj esetében van egy hőmérsékleti tartomány, amelyen belül az életfolyamatok megfelelően le tudnak játszódni. Ezt a tartományt minimum, optimum és maximum értékekkel jellemezhetjük. Az optimumtól általában ±4-5°C-kal lehet eltérni jelentősebb produkcióbeli változás nélkül. Túl alacsony hőmérséklet mellett a növekedés üteme lassul majd megáll. Túl magas hőmérséklet esetén pedig felgyorsul, de a felépítő folyamatok mellett a légzés mértéke egyre nő, míg egy ponton túl a növény több energiát fog felhasználni, mint amennyit megtermel, így a növény végül legyengül. A folyamatosan vegetáló fajok hőmérsékleti igénye 25 °C vagy annál is magasabb, míg a szakaszosan vegetáló növények esetében kettébonthatjuk a hőigényt, és beszélhetünk a vegetációs illetve a nyugalmi időszak hőigényéről.
A vegetációs időszak hőigényét a növény származása határozza meg, mérsékelt égövi fajok esetében ez 16-20°C körüli, melegebb égövről származók esetében 25°C vagy afölötti is lehet, hasonlóan a trópusi növények hőigényéhez. A biológiai nulla fok jelzi azt a minimális hőmérsékletet, ahol a növény kihajt, ez sok mérsékelt égövi növénynél 10°C körül alakul. Aktív hőmérsékleti érték a biológiai nulla fok és a tényleges átlaghőmérséklet közötti különbség. A növények általában a hőösszeget, vagyis az aktív hőmérsékleti értékek bizonyos időszak alatti összegét „érzékelik”, és egy adott szint elérésekor tudnak átlépni másik fejlődési fázisba.
A nyugalmi időszak hőigénye függ annak típusától (téli vagy nyári), és a növényfaj származásától. A téli nyugalmat igénylő növények esetében a hideg periódus szükséges a vegetáció újrakezdéséhez vagy a virágképződéshez – ezt jarovizációs vagy vernalizációs időszaknak nevezzük. Nemcsak a hőmérséklete számít, hanem a periódus hossza is meghatározó lehet az ezt igénylő növények számára. A nyári nyugalom optimális hőmérséklete általában a vegetációs hőmérséklet, 20-25°C körül alakul, és ennek is fontos szerepe lehet virágképződés befolyásolásában, leginkább a hagymás növények esetén használjuk ki ennek a lehetőségét. A valódi nyugalmi időszakot sokszor kényszernyugalmi periódus követi, mely során a növény már elérte azt az élettani állapotot, hogy kész a kihajtásra, de a külső körülmények, például a túl alacsony hőmérséklet, vagy a nagy szárazság további nyugalomban tartja a növényt, és az csak a megfelelő környezeti feltételek beállása esetén fog ténylegesen kilépni a nyugalmi állapotból. Ez kiváló lehetőséget biztosít a kertészek számára egyes hagymás növények virágzásának időzítésében, amiről a későbbiekben részletesen szólunk.
Fontos szerepe van még a növények fejlődésében a cirkadián hőmérsékleti ritmusnak, vagyis a nappali-éjszakai hőmérséklet változásának. A termesztésben csak DIF-ként emlegetett paraméter a nappali és éjszakai hőmérséklet különbségét jelenti. Ha ez az érték pozitív, vagyis a nappalok melegebbek, mint az éjszakák, akkor a növény szármegnyúlással válaszol, fordított esetben pedig nem nyúlik meg a szára. Minél pozitívabb a DIF érték, annál nagyobb lesz a szármegnyúlás, és minél jobban közelít nullához, vagy negatív lesz, annál kisebb lesz a hosszanti növekedése a növénynek. Nem minden növény reagál azonban ugyanúgy a DIF változására, elsősorban a mérsékelt égöviek mutatnak erősebb reakciót, míg a trópusi származásúak kevésbé. Ez érthető is, ha megvizsgáljuk ezen növények élőhelyének klímáját.
Disz.3.07 ANIMÁCIÓ a DIF hatása a szárhosszra
Fény
A növényekben a fénykitettség hatására háromféle folyamat, válaszreakció játszódik le: a fotoszintézis, a fotomorfogenezis és a fototropizmus. A fotoszintézissel, mint energiabeépítő, asszimiláló folyamattal részletesebben most nem foglalkozunk, ellenben a fotomorfogenezisnek, vagyis a fény indukálta egyedfejlődési folyamatoknak, és kisebb mértékben a fototropizmusnak, avagy a fényinger hatására kiváltott helyzetváltoztató mozgásnak fontos szerep jut a növény növekedésszabályozásában. A növények a fény háromféle tulajdonságát képesek érzékelni: a fény hullámhosszát, a fény intenzitását, és a fénykitettség idejét, azaz a fotóperiódust. A növények által hasznosított hullámhossztartomány nagyjából egybeesik az emberi szem számára látható fény tartományával, de ezen tartományon belül nem egyenletes az egyes színekhez tartozó altartományok növényi hasznosíthatósága. A fotoszintézis során a klorofill molekula a vörös tartományt tudja hasznosítani (kis mértékben a kéket is, 440-470 nm között), míg a flavin és karotinoid színanyagok a kék tartományt. A sárga és zöld tartománynak kisebb a jelentősége, de némi élettani hatással ezek is bírnak. A fotoszintézisben hasznosított vörös fény rendelkezik még az alábbi élettani hatásokkal is: serkenti a magcsírázást, a szármegnyúlást és a szárazanyagfelhalmozódást, fokozza az antociánképződést, és elősegíti a generatív életszakaszba történő átlépést. A hormonháztartásra gyakorolt hatását illetően elmondható, hogy eszenciális az organogenezis indukciójához, mert segíti a szövetekben a citokininek felhalmozódását, ezáltal serkenti a hajtások számának növekedését, ugyanakkor csökkenti az auxinok mennyiségét. Infravörös tartományú sugárzás hatására (735 nm felett) a növények különösen erős szármegnyúlással reagálnak. A kék fény ezzel szemben csökkenti a szárhosszúságot, ezáltal erősíti a szárat, növeli annak ellenállóképességét, valamint javítja a virág és a levél színét. Kék fény hatására a növényben nő a gibberellinek és abszcizinsav bioszintézise. A zöld és sárga tartomány hatása gyengébb, de kimutatható, hogy az auxinok lebomlása magas ilyen tartományú fénnyel történő megvilágítás esetén, és emiatt relatív citokinintúlsúly alakul ki a szövetekben, így jó hatással van a járulékos hajtások indukciójára, a rügyek fejlődésére.
A fény intenzitásának és a napi megvilágítás hosszának változása a növények számára jelzi az évszakok múlását, így lehetővé válik, hogy a növények életritmusa alkalmazkodjon a környezeti tényezők periodikus változásához. Fotoperiodikus reakciónak nevezzük a növény válaszát a nappal/éjszaka időtartamának változásaira, mely legfőképp a virágzás megindulásában nyilvánul meg. A fotoperiodikus reakcióidő az a fajra, fajtára jellemző időtartamot jelenti, amíg a növény a fotoperiodikus inger kezdetétől eljut a teljes kivirágzásig. Hosszát hetekben szoktuk megadni. Olykor előfordul, hogy a fényintenzitás változása is befolyásolja a reakcióidő hosszát, tehát egyes esetekben fényösszegről is lehet beszélni, hasonlóan a hőösszeg fogalmához. Ez azonban nem általánosan érvényes, a legtöbb növény esetében a fokozott fényintenzitás nem pótolja a megvilágítottsági tartam rövidülését. A kritikus nappalhossz jelzi azt az időtartamot, ami alatt vagy felett egy adott növényfaj egyedei virágozni kezdenek. A kertészeti szakzsargon rövid nappalnak nevezi a kritikus nappalhossz alatti értékeket, míg hosszú nappal az, ami fölötte van (10-15 óra között szokták megadni az értékeket). Rövidnappalosnak nevezünk egy dísznövényt, ha a számára kritikus nappalhossznál rövidebb nappalok hatására kezd el virágozni, ellenkező esetben pedig hosszúnappalos. A természetben élő növények között a rövidnappalosok ősztől nyílnak, míg a hosszúnappalosok a nyári félév során. Igazából nem a nappal hossza, hanem az éjszakáé a meghatározó a növények számára; ezt bizonyítja, hogy rövidnappalos növény esetén a hosszú éjszaka virágindukáló hatását meg lehet szakítani minimális mesterséges megvilágítással, amiből néhány percnyi elég ahhoz, hogy megszüntesse a hosszú éjszaka hatását. Meg kell még említeni, hogy léteznek nappalhossz közömbös növények is, melyeknél nem függ a virágzás ideje a változó nappalok hosszától, valamint az első két csoportba tartozó növények között is megkülönböztethetünk obligát (minőségileg reagáló) és fakultatív (mennyiségileg reagáló) növényeket. Obligát reakciójú növények csak és kizárólag akkor kezdenek virágozni, ha megfelelő ingert kapnak a sötétperiódus hosszától, míg a fakultatív reakciójú növények az indukciót ki nem váltó nappalhossz mellett is virágozni kezdenek előbb-utóbb, de sokkal vontatottabban, mint ha indukciót kiváltó nappalhossz mellett nevelődnének.
A fotoperiodikus reakciójuk alapján csoportosítható dísznövények néhány példával
-
obligát hosszú-nappalos
fakultatív hosszú-nappalos
fakultatív rövid-nappalos
obligát rövid-nappalos
nappalhossz közömbös
Vágottvirág, cserepes virágos növények
vágott szegfű, Eustoma grandiflorum
Begonia elatior hibridek
Chrysanthemum, Kalanchoë, Euphorbia pulcherrima
Gerbera, Rosa, Strelitzia, Freesia, Pelargonium, Saintpaulia, Primula
Egynyáriak
Centaurea, Linum, Gazania, Lavatera,, Lobelia, Mimulus, Oenothera, Begonia tuberhybrida, Fuchsia hibridek, Rudbeckia
Ocimum, Viola, Salpiglossis, Ageratum, Calendula, Dianthus chinensis, Tithonia, Petunia Grandiflora típus, Salvia farinacea
Celosia, Sanvitalia, Zinnia elegans, Cosmos, Gomphrena, Ipomoea, Tagetes tenuifolia
Tagetes erecta, Lablab purpureus, Ipomoea lobata
Amaranthus, Cleome, Tagetes patula, Impatiens New-Guinea hibridek, Impatiens balsamina, Begonia semperflorens, Zinnia angustifolia
A fejlődés és virágképződés során szerepet játszó kémiai anyagok a növényben
A magasabbrendű növényeknél a sejtek közötti kommunikáció kémiai hírvivők segítségével történik, csakúgy, mint az állati szervezetben. A növényi hormonok ezen kémiai hírvivő anyagok bizonyos típusai, tágabb értelemben véve növekedésszabályozó anyagok. A hormonok hatásmechanizmusára jellemző, hogy speciális fehérjékhez, a receptorokhoz kapcsolódva fejtik ki hatásukat. Nagyon alacsony koncentrációban fordulnak elő, de hatásuk függ a koncentráció nagyságától, valamint keletkezésük helyétől távolabb fejtik ki hatásukat (kivéve az etilént, ami minden sejtben keletkezhet, és hatását helyben fejti ki). Elsősorban a növekedésre és fejlődésre hatnak, és egy jelenséget több hormon is szabályozhat, vagyis nem csak a koncentrációjuk, de az egymáshoz viszonyított arányuk is meghatározó jellegű a hatás kialakulásában. A hormonok gátló és serkentő szerepet is betöltenek. Hagyományosan öt hormontípust lehet elkülöníteni: az auxinok, a gibberellinek, a citokininek, az etilén és az abszcizinsav.
Auxinok
Az indol-3-ecetsav (IES, angol rövidítése IAA) az a vegyület, ami a magasabbrendű növényekben általánosan betölti az auxin szerepét in vivo. Más indolvegyületek (indol-3-vajsav vagy IVS, 4-klór-IES, OH-IES) is rendelkeznek a növényben az IES-hoz hasonló hatással, ám ezek rendszerint az IES előanyagai vagy származékai, és a növényi szervezetben ezek mind IES-vá alakulnak át/vissza. Léteznek azonban szintetikus auxinok is, melyek auxinszerű hatással rendelkeznek, de nem metabolizálódnak IES-vá. Mivel ezen anyagok bomlástermékei is rendelkezhetnek auxinszerű hatással, továbbá ellenállóbbak az enzimes bontással szemben, ezért hatásukat tovább fejtik ki a növényben. Kertészeti szempontból ezek felhasználása jobb eredményeket ad, mint az IES-é, amely egyébként is egy fényre bomló vegyület, és nehéz tárolni, felhasználni emiatt.
Jelentősebb szintetikus auxinok:
- 2,4-diklórfenoxiecetsav (2,4-D),
- 2,4,5-triklórfenoxiecetsav (2,4,5-T),
- 1-naftilecetsav (NES),
- 2-metil-4-klórfenoxiecetsav (MCPA),
- 4-amino-3,5,6-triklórpikolinsav (pikloram),
- 2-metoxi-3,6-diklórbenzoesav (dicamba).
Az auxin fiziológiai hatásait a következőkben lehet összefoglalni:
- a sejtek és szervek, elsősorban a szár megnyúlásos növekedését serkenti,
- hajtás és levéldugványoknál a gyökérkialakulás elősegítése, oldalgyökér fejlődés,
- meghatározó szerepet játszik az apikális dominancia kialakításában,
- szabályozza a levél és termésleválást, a terméskötést, -növekedést, és –érést
- más hormonokkal interakcióban is hat: citokininekkel együtt serkenti a sejtosztódást,
- gibberellinekkel együttműködve pedig a virágok nemi jellegét képes befolyásolni (magas auxin/gibberellin arány esetén nőivarú virágok, ellenkező esetben a hímvirágok kialakulásának kedvez).
Az auxin a növényi szervezetben a növekvő hajtáscsúcsban és a levelekben termelődik, és bazipetálisan képes transzlokálódni a szár háncstestében.
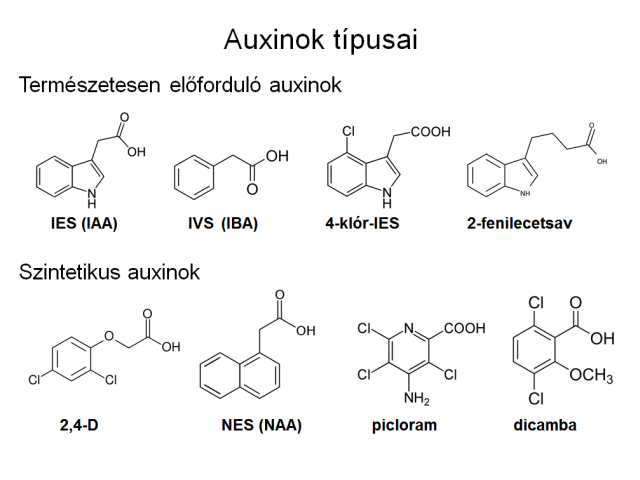
3.11. ábra A természetes auxin, származékai és a mesterséges auxinhatású vegyületek főbb formái
Gibberellinek
A gibberellinek nem csak a magasabb rendű növényekben, de az alacsonyabb rendűekben, baktériumokban, algákban, gombákban is szintúgy előfordulnak. Szerkezetüket tekintve tetraciklusos diterpének, jelenleg több, mint 100 féle gibberellint ismerünk. A növény hajtáscsúcsában szintetizálódnak, és mind a xilémben, mind a floémben képesek transzlokálódni, minden növényi részben előfordulnak. Hatásuk némi átfedést mutat az auxinéval:
- a szármegnyúlás serkentése,
- partenokarpia indukciója,
- apikális dominancia erősítése,
de specifikus hatásaik is vannak, melyeket az auxin nem képes kiváltani, ezek:
- a genetikai és fiziológiai törpeség normalizálása,
- fény- és hidegigényes magvak csírázásának indukciója,
- rügyek nyugalmi állapotának megszakítása,
- a virágzás elősegítése, kiváltása nem induktív környezetben, hideghatást vagy hosszúnappalt igénylő növényeknél,
- a virágok nemi jellegének megváltoztatása a hím jelleg felé.
A kertészeti gyakorlatban a legáltalánosabban elterjedt gibberellin a GS3 (angolul GA3), permetezéssel kijuttatva használják fel az alábbi célok elérésére: szőlőnél a partenokarpia indukciója, a mag nélküli fajták alacsony terméshozamának növelésére, a virágzás kiváltása nem induktív körülmények között, áztatással pedig a magvak csírázásának megkönnyítésére. A virágzás kiváltása csak azon hosszúnappalt igénylő növények esetén lehetséges gibberellin segítségével, melyek rövidnappalos viszonyok között levélrozettát fejlesztenek, és a hosszúnappalok hatására történik a szármegnyúlás és a virágképződés. Ezeknél a növényeknél a gibberellin tehát a hosszúnappalos kezelést képes helyettesíteni. Levélrozettát nem képző hosszúnappalos növények esetén, mint pl a fukszia, a gibberellin nem segíti elő a virágzást, rövidnappalos növényekre pedig szintén nincs hatással sem pozitív, sem negatív irányban. A hideghatás helyettesítésére is alkalmas lehet a gibberellinkezelés, de nem általánosan sikerült bizonyítani ezt a hatást, vannak olyan, a virágzáshoz vernalizációt igénylő fajok, melyeknél nem sikerült a gibberellinekkel virágzást kiváltó hatást elérni. Hagymás nőszirom esetében viszont a GS3 kezelés megfelelően helyettesíti a vernalizációt, abban az esetben, ha a GS3 kezelést követően a növények megkapják a kellő megvilágítási időtartamot a virágképződéshez (ez akkor is szükséges, ha nincs GS 3 kezelés, csak hidegkezelés).
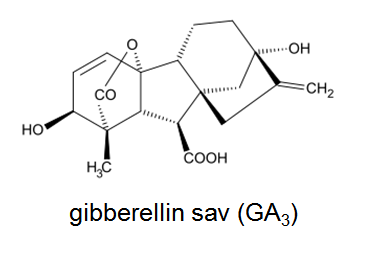
3.12. ábra A gibberellinsav szerkezete
Citokininek
Hasonlóan a gibberellinekhez a citokininek is előfordulnak mind az alacsonyabb, mind a magasabb rendű növényekben, emellett gombákban, baktériumokban. Általánosan elterjedt mivoltukat mutatja, hogy a baktériumokban termelődött citokininek azonosak a magasabbrendűekével, illetve azok analógjai. A gyűjtőnév purinvázas vegyületeket takar, melyek sejtosztódást indukálnak, jellemző rájuk, hogy auxin jelenlétében fenntartják a dohány kallusz tenyészetek folyamatos növekedését. Természetesen is többféle citokinin fordul elő a növényekben, de szintetikusan előállított citokininből is ismerünk jónéhányat. Természetesen előforduló a magasabbrendű növényekben a zeatin, izopentenil-adenin, Populus fajokra jellemző a 6-benzilaminopurin és hidroxilált formái (orto- és meta-topolin). A 6-benzilaminopurint (vagy 6-benziladenin, BAP) szintetikus citokininként is nyilvántartják, csakúgy, mint a kinetint. A növényben a gyökércsúcs termeli a citokinineket, és a xilémben transzportálódnak, a transzspirációs árammal jutnak el a növény föld feletti részeibe.
A fiziológiai hatásuk sok területre terjed ki:
- résztvesznek a sejtosztódás szabályozásában, növekedési, fejlődési és differenciálódási folyamatokban,
- serkentik a sejtmegnyúlást,
- a csírázáshoz vörös fényt igénylő magvak csírázást fényhatás nélkül is megindítják,
- néhány, a virágzáshoz hideghatást igénylő faj esetében a citokininkezelés helyettesíti a vernalizációt,
- késleltetik a levelek öregedését,
- serkentik a tápanyagok mobilizálását,
- az apikális dominancia kialakítása során az auxin antagonistájaként működnek,
- a rügynövekedést serkentik,
- elősegítik a gumók képződését,
- serkentik a kloroplasztiszok érését,
- indukálják enzimek szintézisét.
Kertészeti felhasználásuk a mikroszaporításon kívül fás növények elágazódásának elősegítésében rejlik (benzil-adenin tartalmú készítmények), illetve vágott virágok posztharveszt kezelési eljárása, hogy a vázavízben oldott citokininek hatására lassul a virág hervadása, zöldebbek maradnak a levelek, megnő a vázaélettartama.
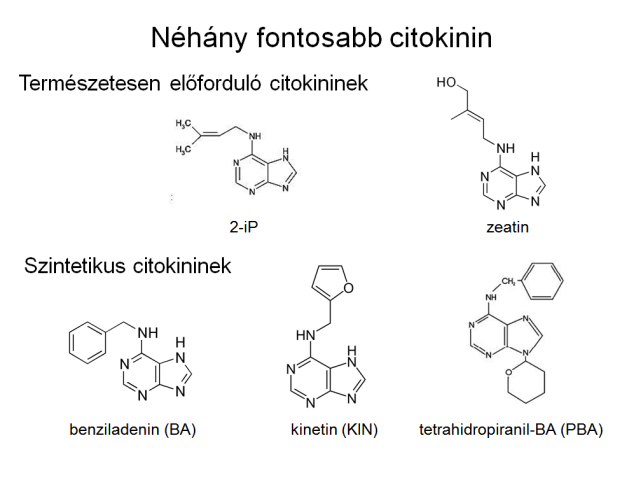
3.13. ábra Néhány fontosabb citokinin hatású vegyület szerkezete
Abszcizinsav
Az 1960-as években több növényből azonosítottak egyszerre egy növekedésszabályozó anyagot, mely ugyanannak bizonyult, és mivel feltételezték, hogy ez a vegyület okozza a növényi szervek leválását (abszcisszióját), abszcizinsavnak (ABS) nevezték el. Ma már tudjuk, hogy ezért az etilén a fő felelős, de az elnevezés megmaradt. A magasabbrendű növényekben a fejlődési fázisok mindegyikében megtalálható vegyület, bár igen kis mennyiségben. Az előbbiektől eltérően ez egy inhibitor (gátló hatású) szabályozó anyag, de egyenlő jelentőségű a korábban tárgyalt hormoncsoportokkal. Fiziológiai hatásai:
- részt vesz a levél leválásában, a leválási zóna kialakulásában
- részt vesz a rügyek nyugalmi állapotának kialakításában és megszűntetésében, más növekedésszabályozókkal összhangban
- szerepe van a magvak nyugalmi állapotának kialakulásában és megszűnésében, külsőleges kezelés esetén csírázásgátló hatást fejt ki
- sietteti az öregedés folyamatát
- serkenti a járulékos gyökérképződést
- gátolja a megnyúlásos növekedést
- virágzást indukál egyes rövidnappalos növényeknél nem induktív körülmények között (szamóca, feketeribiszke)
- vízhiány esetén megnő a koncentrációja, fokozza a gyökerek vízfelvételét és csökkenti a transzspirációt a sztómák záródásának elősegítésével
Hatásai alapján sok felhasználási lehetőség nyílna ABS-val a kertészeti gyakorlatban, de mivel a növények gyorsan metabolizálják az ABS-t, ezért többször és nagyobb koncentrációban lenne szükséges kijuttatni, amit előállítási költsége nem tesz rentábilissé.
Etilén
Az öregedés hormonjaként, stresszhormonként is nevezik az etilént. A növény minden sejtje képes a termelésére, főleg abiotikus stresszorok hatására: hideg, szárazság, sebzés, levegőhiány a gyökereknél, magas ózon és nehézfémkoncentráció. Az öregedő szervek, termések nagyobb koncentrációban termelik, mint az érett, vegetatív szövetek. Fiziológiai hatásai:
- gátolja a megnyúlásos növekedést
- fokozza a hajtás gibberellinérzékenységét
- serkenti a levél- és termésleválást
- gyökeresedést indukál
- megtöri a rügyek és a magvak nyugalmi állapotát
- serkenti a virágkezdemények képződését, a terméskötést, a termés növekedését
- virágoknál az ivari jelleget a nőivarúság felé tolja el
- serkenti a gumóképződést
- elősegíti az öregedési folyamatokat
Kertészeti felhasználása a hatásai miatt jelentős mértékű, de mivel gázról van szó, ezért könnyen elillana, így nem közvetlen formában permeteznek vele, hanem olyan vegyületeket alkalmaznak, amelyekből bomlás során szabadul fel az etilén, pl 2-klóretil-foszfonsav hatóanyagot (Ethrel, CEPA). Mivel a vágott virágok elnyílását gyorsítja, ezért azok tárolóhelyiségében a folyamatosan termelődő etilén koncentrációját csökkenteni kell annak megkötésével. Erre a célra egy porózus – nagy felületű – anyagból készült szűrőt alkalmaznak, melyet kálium-permanganát oldattal itatnak át. Broméliák esetében azonban az idő előtti virágzásindukció fontos eszköze.
Egyéb növekedésgátló anyagok, törpítőszerek
A dísznövénytermesztésben elterjedten használják a növekedésgátló anyagok egy speciális csoportját, az úgynevezett törpítőszereket. Ezek klasszikus értelemben nem tartoznak a növényi hormonok közé, szintetikus vegyületek. A piac igényli a kompakt habitusú, kis méretű növényeket, melyek előállítását sokszor megfelelő genotípusú fajta hiányában csak kémiai kezeléssel lehet megoldani. Erre a célra alkalmazhatók azok a szerek, melyek a szár/szártagmegnyúlást előidéző gibberellinek antagonistájaként működnek, sokszor a gibberellin bioszintézisét gátolják, vagy a csúcsdominanciát blokkolva bokros habitust adnak a növénynek. Ezek a szerek a virágbimbók számát is növelik, viszont a virágzást 1-2 héttel is késleltethetik. Erre alkalmas hatóanyagok a triazol típusú fungicidek közül a paclobutrazol, ancymidol, flurprimidol, illetve a klór-kolin-klorid (CCC), daminozid. A szigorodó környezetvédelmi rendelkezések miatt azonban egyre csökken ezen hatóanyagokat tartalmazó elérhető szerek száma.
Gyakorlati magvalósítási lehetőségek
Szaporítási időpont megválasztása
Akkor van lehetőség kizárólag a szaporítási időpont megválasztásával szabályozni a virágzást, amikor az csak egy megfelelő növényméret/eltelt idő után következik be. Előkezelt (hőkezelt) szaporítóanyag esetén például az Hippeastrum jó példa erre. A legtöbb kultúra esetében viszont az egyéb növekedésszabályozási lehetőségekkel kombináltan használják a szaporítási időpont megválasztását.
Mechanikai kezelések
Mechanikai kezelés alatt egyrészt a növények fizikai ingerlését, másrészt a növény részeinek eltávolítását értjük. A fizikai ingerlés a növényállomány „söpréséből” áll, melyet az állomány fölé rögzített műanyag vagy fa léccel, esetleg kefékkel oldanak meg. A növények hajtáscsúcsát naponta többször, 30-40-szer végigsöprik, aminek a hatása hasonló a természetben a szél hajlítgató hatásához: a növény hosszanti növekedése lassul. Kizárólag egészséges növényeknél szabad használni, és vigyázva, hogy ne okozzunk sérülést. Magyarországon nem nagyon alkalmazzák a módszert, de mivel beruházásigénye alacsony, és a környezetre káros vegyszereket sem igényel, elterjedése várható a jövőben.
A fizikai ingerlésnél erősebb fiziológiai hatással bír a hajtáshelyzet megváltoztatása, a lehajtogatás módszere, amit növényházi vágott rózsa termesztés esetén gyakran alkalmaznak. A hajtáshelyzet erőteljes megváltoztatását a feltörekvő szárak legalsó pár rügy környékén történő megtörésével-megcsavarásával és lehajlításával végzik. Ennek eredménye a növényben az apikális dominancia megszűnése, és így a hajlítás környékén a rejtett rügyek kihajtásának elősegítése. Szintén az apikális dominancia megszüntetését használja ki a különböző magasságokban elvégzett visszacsípés, visszavágás, visszatörés. Optimális hatásra akkor számíthatunk, ha a hajtáscsúcs mellett a legfiatalabb 1-2 levelet is eltávolítjuk a növényről, ilyenkor még a kihajtás kellően gyors a levélhónalji rügyekből, és jobb bokrosodást is elérhetünk, mintha csak a hajtáscsúcsot távolítjuk el. Idősebb részekig történő visszavágás, -törést ritkábban alkalmazunk, főleg akkor, ha elkéstünk a megfelelő magasságban történő visszacsípéssel. Ilyenkor a levélhónalji rügyek kihajtása vontatottabb, és kevesebb is hajt ki. Bár nem kifejezetten kezelésnek minősül, mégis ebben a pontban tárgyalható még a cserépméret és –forma hatása a növekedésre.

3.15. ábra A visszacsípés hatására kihajtó oldalrügyek hajtásokat fejlesztenek és bokrosítják a mikulásvirágot
A termesztőedény tulajdonságai, elsősorban a térfogata adott maximális gyökértömeg létrejöttét határozza meg. Egy adott gyökértömeg pedig meghatározott föld feletti részt képes tápanyaggal és vízzel ellátni, tehát a növény potenciális mérete eleve korlátozva lesz. A cserépméret megválasztása során azonban arra is kell ügyelni, hogy a maximális méretet elért növény egy idő után könnyen elöregszik az adott méretű edényben tartva.
Rövidnappalos kezelés
Kivitelezése elsötétítéssel történik, Magyarországon áprilistól szeptemberig lehet rá szükség. A növényállományt egy fényt át nem eresztő anyaggal takarják le. Általában sűrűn szőtt anyagot használnak, melyet a növények fölött kialakított vázrendszerre húznak ki. A fekete fólia is alkalmazható erre a célra, de figyelembe kell venni, hogy nagyon bemelegedhet alatta az állomány, mivel nem szellőzik. Ennek kiküszöbölésére találták ki azt a fóliát, melynek külső oldalát vékony rétegben alumíniummal vonják be. Ez visszaveri a hőt, megakadályozza a túlzott felmelegedést. Megfelelő vázrendszer esetén a takaróanyag energiaernyőként is alkalmazható a fűtési szezonban a költségek csökkentésére.

3.16. ábra A korallvirág termesztése során a rövidnappalos kezelést az állomány fölé kifeszített fekete függönnyel valósítják meg

3.17. ábra A rövidnappalos kezelés másik megvalósítása: az állomány fölé helyezett alagútvázra fényt át nem eresztő fóliát húznak később
Asszimilációs pótmegvilágítás
Fényigényes kultúráknál, vagy fakultatív hosszúnappalos növények esetén alkalmazzuk a nagyobb hozam elérésére (például rózsa, liliom). A legtöbb dísznövénynél az asszimilációs pótmegvilágítás javasolt erőssége minimum 4000-5000 lux. A hosszabb időn át biztosított alacsonyabb fényintenzitás hatásosabb, mint a rövid ideig adott intenzívebb megvilágítás – pl a 24 órán át adott 4000 lux jobb hatást ad, mint a 12 órás 8000 luxos pótmegvilágítás. Magyarországon október végétől március elejéig alkalmazzák, napfényes időben 4-6 órával hosszabbítjuk meg a nappalt, borús idő esetén akár egész nap, 24 órán át is mehet a megvilágítás a növényeknek. Hosszúnappalos inger biztosításához elegendő az állomány fölé lógatott 60-100 wattos villanykörte is, asszimilációs pótmegvilágításhoz viszont elengedhetetlen a nagyobb fényintenzitást, és a növények számára jobb spektrumot biztosító HPS (nagynyomású nátriumgőz-lámpák) lámpák alkalmazása. Ezekből a 400 wattos izzók terjedtek el széleskörben, ugyanis ezek hőtermelése még nem olyan nagy, hogy a növények fölé 3-4 méter magasságba elhelyezett izzók hője károsítaná a növényállományt. Ezeket 16 négyzetméterenként elhelyezve (vagyis egy fényforrás fénye 4 m magasságból 16 négyzetméterre vetül), az általában 50000 lument biztosító 400 wattos izzó a növényállomány szintjében 50000/16=3125 luxnak megfelelő megvilágítást ad.

3.18. ábra Az asszimilációs pótmegvilágítást biztosító nagy nyomású nátriumgőz-lámpák
A jövőben alkalmazásra kerülhetnek növényházakban is LED-ekből álló megvilágítási rendszerek, melyeket a mikroszaporításban már kezdenek egyre több helyen alkalmazni. Nagy előnye a LED-es rendszereknek, hogy szinte tetszés szerint választható a kibocsátott hullámhossztartomány, és így könnyen állítható a kék/vörös/távoli vörös tartományok aránya a sugárzott fényben. Másik jövőbeli lehetőség még a fénnyel történő növekedésszabályozásra a fényszűrők használata, mellyel az üvegházba belépő természetes fényben a termesztési program aktuális szakaszának megfelelően adott hullámhossztartomány-arányokat állítunk elő.
Hőkezelések
A hőmérséklet változtatásával is elérhetünk növekedésszabályozást a növényeknél. A korábban említett DIF értékének minél kisebbre csökkentésével például növekedésgátlást érhetünk el, és csökkenthető a kémiai retardánsok felhasznált mennyisége. A DIF szabályozásán túl az éjszakai hőmérséklethez képest hajnalban 3-6°C-kal alacsonyabb hőmérséklet tartása 2-3 órán keresztül hasonló, vagy akár jobb eredményt is okozhat a növekedés visszatartásában. A DIF kezelést nem szükséges az egész termesztési ciklus alatt tartani, elegendő csak a legaktívabb vegetatív növekedés periódusában alkalmazni. Általánosságban elmondható, hogy a DIF csökkentésével megnő a kultúraidő (ha a DIF csökkentését a napi átlaghőmérséklet csökkentésével érjük el – vagyis nappal és éjszaka egyaránt kevesebbet fűtünk).
A virágzáshoz hidegperiódust igénylő fajok esetében lehetőség van a nyugalmi időszak hosszát befolyásolni a megfelelő hőmérsékleten való tartással. Ezeknek a fajoknak sokszor rövidebb a hidegperiódus igénye, mint a természetben előforduló tényleges hidegperiódus hossza, és az aktív nyugalmuk lejárata után már csak a még fennálló hideg időjárás miatt nem kezdenek el hajtani. A termesztés során ezeknél a fajoknál lehetőség nyílik a hidegperiódust lerövidíteni arra a minimális időtartamra, amíg a növényben lezajlik az aktív nyugalmi időszak, ezután a hőmérséklet emelésével pedig virágoztatásra késztethetők, korábban, mint ahogy a természetben tennék azt. A kihajtás gátlását, időzítését pedig a meghosszabbított hidegperiódussal érhetjük el, vagyis a kívánt kihajtás idejéig hidegben tartjuk a növényt annak ellenére, hogy már lezajlott benne az aktív nyugalom (például a „2 fokos” tulipánhagyma, amely megkapta a vernalizáció mindegyik szakaszát, és ezután a kiültetésig 2 °C-on tárolják). Mérsékelt égövön célszerű az ilyen növények termesztését úgy időzíteni, hogy a hidegkezelés periódusa az egyébként is hideg évszakra, télre essen (flamingóvirág, húsvétikaktusz). A hidegkezelés alatti fényellátás mértéke befolyásolhatja a hidegkezelés hosszát – a gyengébb megvilágítás meghosszabbítja, a jó fényellátás pedig megrövidíti annak tartamát. Egyes szubtrópusi fajok (például a Cymbidium) olyan erős fényintenzitást is igényelhetnek a hidegperiódus alatt, amit nálunk a téli évszak során a borús időjárás miatt nem tudunk számukra biztosítani (asszimilációs pótmegvilágítás nélkül), így ezeknél a növényeknél a nyári szakaszban kell a hidegperiódust megadni az üvegház hűtésével, ami technikailag igencsak költséges, de megvalósítható. A legelterjedtebb megoldás a hűtőfalas üvegház, ahol az egyik oldalfalon vizet folyatnak le egy nagy felületet biztosító anyagon, míg a másik oldalon elhelyezett ventilátorokkal keresztülszívatják a levegőt a termesztőházon.

3.19. ábra A hűtőfalas üvegház vizet csordogáltató oldala kívülről: az árnyékolással óvják a fal felmelegedését

3.20. ábra A hűtőfalas üvegház ventilátoros oldala
A víz evaporációjának hatására a hőmérséklet így csökkenthető. Melegkezelésről is beszélhetünk néhány növény esetében. Holland nősziromnál például a nyári kényszernyugalmi periódust 30 fokon való tartással lehet imitálni, és így akár egy egész évig lehet tárolni a hagymáját a kultúraindítás kívánt idejéig. Jácint esetében a nyári kényszernyugalom idején végzett melegkezelés segítségével állítják elő a „szalagosodott” virágokat, vagyis az egybeforrott virágszálakat a minél dekoratívabb hatás elérése végett. Ennek során a hagymáját 10 napig 23°C-on, 2 hétig 30°C-on, majd 3 hétig 25,5°C-on tartják
Kémiai szerek alkalmazása
A kémiai szereket elsősorban a dugványok gyökeresedésének elősegítésére, illetve a növény törpítésére alkalmazzuk. A hormonok hatásának ismertetésekor már említettük, hogy az auxinokat használják a gyökeresedés kiváltására. Ezek közül is elsősorban a szintetikus auxinokat, az indol-3-vajsavat (IVS) és a naftil-ecetsavat (NES). Előnyük, hogy jóval stabilabbak a természetes auxinnál, nem bomlanak hő és fény hatására. Vízben csak a nátrium- és káliumsóik oldódnak, ezért vizes oldat készítéséhez csak ezeket használhatjuk, egyébként a por alakban forgalmazott tiszta formájuk csak szerves oldószerekben, pl etil-alkoholban oldódik jól. A dugványok kezelése történhet 50%-os etil-alkohollal elkészített 0,1-0,5%-os oldattal, ilyenkor a dugványok talpát 1-1,5 cm mélyen 5 másodpercre bemártják az oldatba, majd lerázzák és félreteszik néhány percig szikkadni dugványozás előtt. Másik lehetőség az oldat helyett a porkészítmény, ami gyárilag is kapható. A gyökereztető por úgy készül, hogy az auxin tömény etil-alkoholos oldatát talkumporral (magnézium tartalmú szilikátásvány, melyet porrá őrölve töltő- és vivőanyagként alkalmaznak) keverik, és megvárják, amíg az alkohol elpárolog. A talkumpor megszárad, és egyenletesen elosztva tartalmazza a hormont. Ennek felhasználása hasonló, mint az oldaté, csak itt ügyelni kell arra, hogy enyhén nedves legyen a dugvány talpa bemártás előtt, mert csak így tapad rá kellő mennyiségű szer.
A törpítőszerek kijuttatása beöntözéssel és permetezéssel történhet. A beöntözés kisebb szakmai felkészültséget igényel, ugyanakkor több szer fogy. Növényfajonként is eltérő lehet, hogy a beöntözésre vagy a permetezésre reagálnak-e jobban a kezelt növények. Permetezéses kijuttatás után a lemosódás elkerülése végett nem célszerű 24 órán belül párásítani sem. A kijuttatandó anyag töménysége mindig a szer típusától függ, erről a szer gyári tájékoztatója ad felvilágosítást. A legtöbb esetben a kijuttatás után a növények sárgulni kezdenek, ami természetes jelenség, de jól használható egyfajta indikátorként is: jelzi a szer kijuttatásának egyenletességét. A sárgulás megszüntetését nitrogéntartalmú levéltrágyázással lehet elérni, melyet a kezelést követő 2. vagy 3. napon lehet megkezdeni.

3.21. ábra A törpésítőszerrel nem kezelt (0), és kezelt Ismelia carinata növények: Caramba (1), Regalis (2), Alar (3)
Ellenőrző kérdések
- Milyen célt szolgál a dísznövénytermesztésben a mesterséges növekedésszabályozás?
- Milyen fejlődési szakaszok különíthetőek el a növények élete során? Mik a fő jellemzői ezeknek a szakaszoknak?
- Mit nevezünk rejuvenilizációnak, és hol van szerepe a kertészeti termesztés során?
- Milyen környezeti tényezők befolyásolják a növények virágképződését?
- Mit nevezünk vernalizációnak? Mely növényeknek van rá szükségük?
- Mit jelent a biológiai nulla fok?
- Hogyan befolyásolja a DIF érték a növények hosszirányú növekedését?
- Melyik három élettani folyamat során van szerepe a fénynek a növényi szervezetben?
- Milyen biokémiai változásokat indukál a fény vörös és kék tartományba eső része, és hogyan befolyásolja ez a növény morfológiai tulajdonságait?
- Mi a különbség a rövid- és hosszúnappalos növények között?
- Mit takar a fotoperiodikus reakcióidő fogalma?
- Mire lehet használni az etilént, mint növényi hormont a kertészeti gyakorlatban?
- Milyen módon lehet megoldani üvegházban a rövidnappalos kezelést?
- Hogyan lehet Cymbidium-nál virágzásindukáló körülményeket biztosítani mérsékelt égövön üvegházban télen illetve nyáron?
- Mi a törpítőszerek biokémiai hatásmechanizmusa, és hogyan kell kijuttatni őket?
Irodalomjegyzék
Erdei, L. (szerk.) (2004) Növényélettan. Növekedés és fejlődésélettan, JATEPRESS, Szeged
Haraszti, E. (1977) Zsebkönyv a növények életéről, Natura, p. 233-281.
Hortobágyi, T. (szerk.), Simon, T. (szerk.) (2000) Növényföldrajz, társulástan és ökológia, Nemzeti Tankönyvkiadó, Budapest, p. 280-352.
Pethő, M. (2002) Mezőgazdasági növények élettana, Akadémiai Kiadó, Budapest, p. 268-327, p. 374-433
Rünger, W. (1977) Virágképződés és virágfejlődés, Mezőgazdasági Kiadó, Budapest, p. 5-193.
Schmidt, G. (szerk) (2002) Növényházi dísznövények termesztése, Mezőgazda Kiadó, Budapest, p. 84-102.
Turcsányi, G. (szerk.) (2001) Mezőgazdasági növénytan, Mezőgazdasági Szaktudás Kiadó, Budapest, p. 51-173.
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028