Szárazföldi életközösségek
Minden élőlény számára létezik egy olyan hőmérsékleti spektrum (tolarenciaspektrum), amelyhez alkalmazkodott. Az élőlények kétféle módon reagálnak a változó körülményekhez, változtatnak elterjedésükön, illetve fenológiai ciklusukon. A felmelegedéssel együtt az a terület, ahol az élőlény számára megfelelő hőmérséklet uralkodik, a sarkok felé, illetve hegyvidéki területeken a magasabb régiók irányába tolódott el. Számos tanulmány foglalkozott azzal, milyen hatással lesz ez az élővilágra (pl. Thuiller et al. 2004, 2005, Kelly et al. 2008, Eaton 1996).
Kelly és munkatársai (2008) leírták, hogy harminc év leforgása alatt az általuk vizsgált 1350 hegyvidéki növényfaj többsége átlagosan 64,7 m-rel magasabban fordul elő. A növényzeti régiók egységesen, mindenhol ennek megfelelően a magasabb területek felé mozdultak el. A területek alsó határán a növények képtelenek voltak elviselni a megemelkedett hőmérsékletet és az időnként megjelenő szélsőséges időjárást, elpusztultak, ezt követően helyükre más faj lépett. Az okokat vizsgálva megállapították, hogy az átalakulásban döntő szerepet játszott a hóhatár fentebb tolódása, a hőmérséklet emelkedése és az aszályos időszakok megléte. Parmesan és Yohe vizsgálatában (2003) szereplő 99 faj átlag 6,2 km-t vándorol északra évtizedenként. 434 faj a klímaváltozásnak megfelelően változtatta elterjedését. A mérsékelt övi fajok terjednek, de a sarkvidékiek területe zsugorodik.
Az életközösségeket leginkább veszélyeztető probléma, hogy az élőlények nem egyformán reagálnak a megváltozott környezeti körülményekre, ezáltal a táplálékhálózatok számottevő mértékben átalakulhatnak. A tápláléklánc különböző tagjai másként reagálnak a klímaváltozásra (Crick 2004, Harrington et al. 1999), mára azt is tudjuk, hogy egy faj, minél magasabb helyet foglal el a táplálékláncban annál érzékenyebb a klímaváltozásra (Voigt et al. 2003). Külön érdekesség, hogy a közel rokon fajok sem alkalmazkodnak feltétlenül ugyanolyan mértékben (Végvári et al. 2009).
Eltolódott évszakok:
Számos megfigyelés támasztja alá azt, hogy a vegetációs időszak kezdete az elmúlt évtizedekben korábbra tolódott (Parmesan és Yohe 2003, Visser és Both 2005). Észak Amerikában a fagymentes időszak beköszönte az elmúlt 60 év alatt 11 nappal hamarabbra került (Easterling et al. 2000). Ez azt eredményezi, hogy korábban következik be a rügyfakadás, az állatok ezzel párhuzamosan hamarabb jelennek meg. Parmesan és Yohe átfogó tanulmánya (2003) szerint az általuk tanulmányozott 677 faj 62%-a kezdi meg korábban a tavaszi életciklusát.
Egy előrejelzés szerint a madárfajok 75%-a hamarabb kezdi meg szaporodási ciklusát 2080-ra. A korábban fészkelő madarak nagyobb mértékben változtattak a tojásrakásukon, mint a később költők. Ezen felül vannak fajok, amelyek telelőhelyükről hamarabb kerülnek elő, vagy a hibernációból korábban ébrednek fel, amikor még nem minden környezeti feltétel kedvező számukra (pl. vándorrigó (Turdus migratorius), sárgahasú mormota (Marmota flaviventris), stb.) (Crick 2004, Inouye et al. 2000 ).
Már találtak néhány bizonyítékot arra, hogy a hosszú távú vonuló madárfajok kevésbé képesek alkalmazkodni a klímaváltozáshoz rövid távon vonuló társaiknál, főleg azok a fajok, amelyek nem a hőmérséklet változáshoz, hanem valamilyen más ingerhez igazítják a vonulás megkezdésének idejét. Végvári és munkatársai (2009) a madarak érkezési idejét vizsgálták 93 fajon. Azt találták, hogy trendszerűen hamarabb érkeznek vissza, de vannak csoportosulások is: azok a fajok, amelyek hosszú távon vonulnak, kevésbé, míg a rövid távú vonuló fajok jobban alkalmazkodnak a korábban beköszöntő tavaszhoz. Azok a fajok, melyek az evezőtollakat a nyári időszakban váltják, szintén hamarabb érkeznek a telelőhelyen vedlő társaiknál. A változatos táplálékot folytató, sok fiókát felnevelő fajok is hamarabb megérkeznek.
Erre, az eltérő viselkedésre lehet szemléltető példa az örvös légykapó (Ficedula hypoleuca) és széncinege (Parus major) állományának változása (Both et al. 2005). Kiderült, hogy a hosszú távon vonuló madarak, mint például az örvös légykapó kevésbé képesek alkalmazkodni a klímaváltozáshoz, mivel viszonylag rögzített genetikai program szabályozza érkezésük időpontját. A széncinege tojásrakásának idejét pedig az Észak Atlanti Oszcilláció határozza meg (Sanz 2002) . Emiatt, a légykapók, bár kimutatható trend szerint egyre korábbra időzítik érkezési idejüket, mégis késnek: a fő táplálékuk (hernyók) egyedszám csúcsa korábbra tolódik, így a fiókákat nem képesek felnevelni, erős szelekció hat a későn érkezőkre. Ezen túlmenően a hernyók egyedszám csúcsa meghatározza a cinegék másodköltésének meglétét is, ezek a madarak jól alkalmazkodnak ehhez a trendhez, növekszik az állományuk.
A gazda – parazita kapcsolat egyik ékes példája a kakukk (Cuculus canorus) és fészekgazdáinak esete. Möller és társai (2010) egy friss cikkben számolt be arról, hogy a kakuktojással fertőzött fajok közül azok, amelyek hosszú távon vonulnak, nagyobb mértékű parazitáltságot mutatnak, mint a rövid távon vonulók, vagy áttelelő fajok. Ennek oka is a fajok eltérő fenológiai válaszában rejlik. A kakukkok, mivel hosszú távon vonulnak, nem olyan gyorsan alkalmazkodnak a korábbra tolódott vegetációs periódushoz, mint rövid távon vonulók. Így később érkeznek, amikor gazdaállataik akár már fészkelésüket is megkezdték. Ez jelentős hátránynak bizonyul, mivel kisebb eséllyel képesek tojásaikat ezekbe a fészkekbe tojni. Ennek köszönhetően mutatkozik egy évtizedes trend, ami a kakukkfiókák arányát a hosszú távon vonuló madarak felé tolja el.
A növényzet átalakulása:
Összességében elmondható, hogy a legtöbb életközösségben várhatók kritikus változások, amelyek alapvetően megváltoztatják egy terület összképét. Ez az átalakulás a társulás alkotó fajok elterjedésének változásával hozható összefüggésbe. A modellek tanulsága szerint az északi, zömében tűlevelű fafajok elterjedése nagy mértékben visszaszorul, elterjedési területük északi részén maradhatnak meg, vagy menedékhelyekre szorulhatnak. A lombhullató fák aránya ezzel szemben várhatóan nőni fog az északi terjeszkedés révén. Délen ezzel szemben a lombhullató fákat a mediterrán fajok váltják fel, keveredő funkcionalitás jelentkezik, hosszabb vegetációs periódussal, több megkötött szénnel. Még délebbre pedig bokrok és füvek veszik át az az uralmat, de ez a tüzek gyakoriságát is várhatóan növekedni fog (Thuiller et al. 2005).
A magasabb szélességi körök ökoszisztémái is átalakultak az elmúlt évek során: A hőmérséklet emelkedésével egy időben a bozót sűrűsége nőtt, a fák elterjedésének északi határvonala kitolódott. A megváltozó albedónak és energia háztartásnak köszönhetően az elmúlt 25 évben fotoszintetikus aktivitás növekedést figyeltek meg. Ezzel ellentétes hatást fejt ki, hogy a csapadékstressz felülmúlja a meghosszabbodott vegetációs időszak által adott előnyt. Ennek következtében a nettó primer termelés csupán 2000-ig nőtt (1,4%), azóta csökken. A melegebb levegő miatt kevesebb csapadék hull, így a növények számára is kevesebb víz érhető el (Bunn et al 2007).
Egy közösségben minimális hatása van annak, ha egy fajt hasonló funkcióval bíró fajjal helyettesítünk, de ha egy más funkciót betöltő fajjal tesszük meg ezt, nagy változást idézünk elő. Például ha fűfajokat széleslevelű lombhullató fafajokkal helyettesítünk, jelentős mértékben megváltoztathatjuk a mérsékelt övi ökoszisztémák nitrogén és szénforgalmát. Emellet a szoros kapcsolatok révén más fajokra is jelentős hatással van a változtatás (Thuiller et al. 2006.).
Az Amazonas menti erdők területének csökkenésének velejárója, hogy csökken az evapotranspiráció, melegebb, szárazabb klíma alakul ki. Ez azért lényeges, mivel a szárazabb klímán nagyobb esélye van az erdőtüzeknek is. Ezzel együtt csökken a visszatartott CO2 mennyisége is. Dél-Amerika esőerdeit az emberi hatások mellett az átalakuló klíma veszélyezteti. A legveszélyeztetettebb erdőrészek azok, amelyek közel vannak az erdőszegélyekhez. A folyamatot gyorsítja, ha megnyílik a lombozat, mivel több levél hullik le, sok elhalt faanyag keletkezik, amelyet az előforduló tüzet okozó események meggyújthatnak. Ebből olyan avartüzek keletkeznek, amelyek a fiatal fákat pusztítják, esetenként mélyen behatolva az erdőbe. Ezek a tüzek még inkább feldarabolják a lombozatot (Laurance és Williamson 2001).
A globális szén dioxid kibocsájtás 10-15%-a viszont a trópusi erdők degradációjából származik. Asner és társai (2010) műholdas és helyileg felvételezett térképek alapján határozták meg az erdőség által tárolt szén mennyiségét. Nagy hatással van a bányászat, állattenyésztés és infrastrukturális beruházás, szelektív fakitermelés a terület által megtartott szén mennyiségére (fák magasságára). Széles régiónkénti szétdarabolódást találtak alapkőzet szerint az erdőségekben.
Az elmúlt években sok lombhullást és fapusztulást tapasztalnak a szárazság előtti és aszály alatti időszakokban, mikor is a szegélyekben található pionír fajok haltak meg nagyobb számban, így a megújulás esélye jelentős mértékben csökken. A feldarabolódásnak köszönhetően egyre nagyobb területen szárad ki az avar és csökken az erdőség szénbeépítő képessége is. Az erdőpusztulás a csapadék mennyiségét akár 20%-al is csökkentheti (Lean et Warrilow 1989; Shukla et al. 1990) . Az erdőtüzek pedig sokszor a gyújtogatás miatt jönnek létre: 1997-ben 47000 tüzet figyeltek meg egy év alatt az Amazonas medencében, amelyet farmerek okoztak (Brown 1998). Amint az aszályok következtében a mélységbe levő vízmennyiség is lecsökken, akkor ezek az erdők sokkal fogékonyabbak lesznek a tűzre (a szárazabb esőerdők fáinak gyökere képes lehatolni a mélyben levő vízkészletekhez, de ha a mélységi vizek elapadnak, akkor az erdő kiszáradhat). Az erdőtüzek ezen felül jelentős üvegházhatásű gáz fokozó jelenségek (Laurance et Williamson 2001 ; Houghton 1991; Fearnside 2000). Mint látjuk, a folyamatban rengeteg visszacsatolási hurok van, amely aggasztó kilátást nyújt az előttünk álló évtizedekre.
Hazánk szárazföldi vegetációja
Hazánk nemcsak éghajlati, hanem biogeográfiai szempontból is átmeneti területen van: a közép-európai lomberdőövezet és a kontinentális erdőssztyepp határán, jelentős szubmediterrán hatásokkal. A várható felmelegedés hazánk területét több okból is különösen érzékenyen érintheti. Egyrészt a klimatikusan meghatározott növényzeti övek határai a klímaváltozás szempontjából fokozottan érzékenynek számítanak, másrészt a Kárpát-medence elszigetelt helyzete fokozott veszélyeztetettségre hívja fel a figyelmet. Magyarország természeti értékei már ma is nagyon szűk területre szorultak vissza, és veszélyeztetettségük klímaváltozás nélkül is drámai. Ebből következően, hazánkban a klímaváltozás hatása mindenképpen alapvető átstrukturálódásban, az állapotok gyökeres megváltozásában nyilvánulna meg, amely felhívja a figyelmet a természetvédelmi és általánosabb értelemben biodiverzitás-megőrzési alkalmazkodás jelentőségére, ezen feladatok súlyára és fontosságára. (Hufnagel és Sipkay 2012, Harnos et al. 2008)
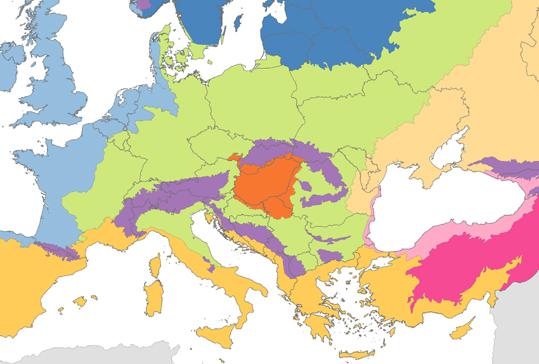

Európa biogeográfiai régiói, 2005. (forrás: EEA)
Magyarország klimatikus viszonyainak az évszázad végére várható, igen jelentős megváltozása a biogeográfiai zónák eltolódását is okozhatja.
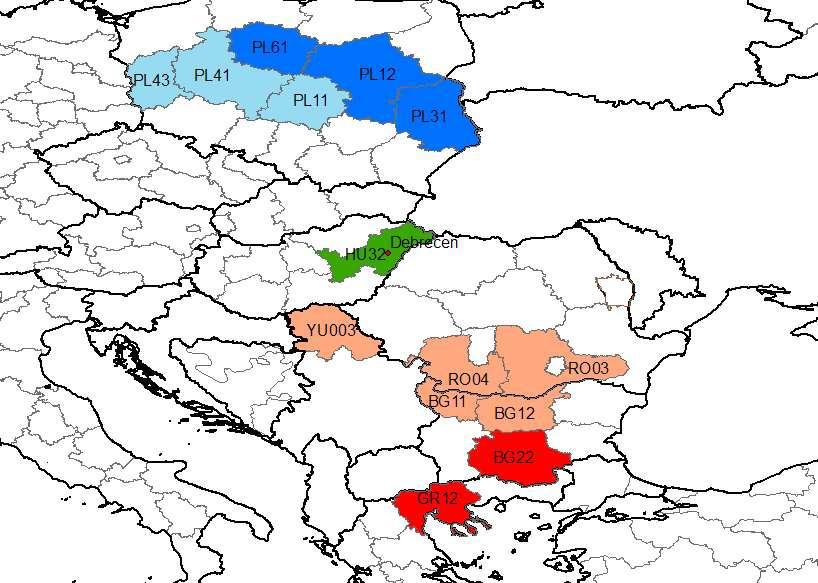
Klímazónák várható eltolódása, földrajzi analógia alapján (Debrecen lehetséges analóg NUTS2 régiói a vizsgált időszakokra)
A megváltozott klímához a növények vagy helyben alkalmazkodnak, amennyiben képesek erre, vagy elvándorolnak, elsősorban szaporító képleteik révén. A terjedési folyamatot viszont nagyban hátráltatja az élőhelyeik emberi tevékenységnek köszönhető feldaraboltsága. A természetes vegetációval borított területek között ugyanis nincs elég kapcsolat, így nem tudnak vándorolni a fajok, és nem találhatják meg azokat a menedékeket, ahol átvészelhetik a számukra kedvezőtlen időszakot.

Dolomit sziklagyep a budaörsi Odvas-hegyen – számos védett faj otthona, a fragmentáció áldozata (fotó: Petrányi Gergely)
A pannon vegetáció legveszélyeztetettebb csoportját az endemikus fajok képezik, mivel teljes elterjedésük csak a Kárpát-medencére korlátozódik, az egész világon csak itt fejlődtek ki. Ilyen fajok például a magyar nőszirom (Iris aphylla subsp. hungarica), a magyarföldi husáng ( Ferula sadleriana) és a pilisi len (Linum dolomiticum). Mivel hazánkban a klímaváltozás leginkább a vizes élőhelyek kiszáradásában vagy a száraz élőhelyek még szárazabbá válásában mutatkozhat meg, így a nedves élőhelyeket kedvelő (atlanti-szubmediterrán, de még inkább szubatlanti) fajok részéről is fokozott reakció várható. A nagy elterjedésű, eurázsiai vagy globális elterjedésű (kozmopolita) fajokat érintheti legkevésbé a klímaváltozás. A délies elterjedésű, szárazságtűrő fajok közül kerülhetnek ki a klímaváltozás haszonélvezői, különösen, ha jó terjedőképességgel rendelkeznek. Azonban e fajok jelentős része inváziós gyomnövény, fokozott terjedésük nagy figyelmet érdemel.

A Pilis–Budai-hegység dolomitszikláinak és a meleg lejtők sztyeppvegetációjának bennszülött növénye, a pilisi len (Linum dolomiticum) (forrás: DINPI)
A biológiai invázió problémája különös jelentőséggel bír a klímaváltozással kapcsolatban. A klímaváltozás hatására a jelenleg is problémát okozó invazív (azaz: özön-) növényfajok hazai elterjedése megváltozhat, de új fajok is megjelenhetnek, vagy jelenleg nem invazív idegen fajok válhatnak özönfajokká. Bár elképzelhető, hogy egyes özönfajok visszaszorulnak a számukra kedvezőtlenné váló klíma miatt, általánosságban elmondható, hogy a gyors környezeti változásokra a gyorsan terjedő fajok képesek jól reagálni, így a sikeres inváziós fajok – mivel ezzel a tulajdonsággal alapvetően rendelkeznek – a jelenleginél is nagyobb arányú térhódítása várható. Mindez nem csupán a hazai flórára jelent nagy veszélyt, hanem számos özönnövény – allergén hatásának következtében – az emberi egészségre is ártalmas lehet.
A klímaváltozásra a növényfajok fenológiai változásokkal is reagálhatnak. Számos vizsgálat alapján a kora tavaszi fajok reagálhatnak a legszembetűnőbben, korábban hajtanak, és korábban virágoznak. Hazánkban is végeztek hosszú távú fenológiai vizsgálatokat. Kiderült például, hogy az akác virágzása az elmúlt 100 évben, régióktól függően mintegy 3-8 nappal előbbre tolódott.

Fehér akác (Robinia pseudoacacia) (forrás: http://hortiplex.gardenweb.com)
A természetes állapotú, nagy fajgazdagsággal bíró közösségek stabilabbak, mint a természetes állapotát tekintve leromlott, vagy másodlagos állományok. A fajgazdagság szerepét időjárási változások okozta kilengések tompításában már számos esetben kimutatták. A fajokban gazdag természetes ökológiai rendszerek a klímaváltozás negatív hatásait is jobban képesek tompítani. Mindez azért is jelentős tény, mert a sűrűn lakott Európában – így hazánkban is – több leromlott állapotú, fajokban elszegényedett élőhely van, mint természetközeli állapotú.

Egyetlen fafaj által uralt alföldi akácültetvény (forrás: http://zoldhullam.blog.fn.hu/))

Természetközeli állapotú keményfa-ligeterdő (http://www.novenyzetiterkep.hu, Bölöni János)
Állatvilág
Az ismert élőlényfajok több mint háromnegyedét gerinctelen állatfajok, többségükben rovarok és más ízeltlábúak alkotják. A rovarok szerepe azonban nemcsak bolygónk biodiverzitásában ilyen kiemelkedő, hanem a természetközeli és mezőgazdasági ökoszisztémák működését is alapvetően befolyásolják. E tekintetben talán elegendő a virágos növények megporzására, a kártevő rovarokra, a biológiai védekezés ágenseire, a talaj folyamatait fenntartó ízeltlábúakra vagy a humán-, állat- és növény-egészségügyi kórokozók vektoraira utalnunk.

Bükköseink látványos lakója a havasi cincér (Rosalia alpina) (fotó: Petrányi Gergely)
A klíma változékonysága alapvető hatással van a rovaregyüttesek szerkezetére, a regionális faunák összetételére, és az egyes lokális élőhelyek (mikrohabitatok, biotópok) szezonális állapotváltozásaira is. Az időjárás közvetlen hatását a fenológiai és populációdinamikai folyamatokon keresztül fejti ki, amely közvetve eredményezi az elterjedési viszonyok és a biodiverzitás átalakulását.
A rovarközösségek azonban nemcsak önmagukban fontosak, hanem (úgy a környezet- és természetvédelem, mint a mezőgazdaság tekintetében) a klímaváltozás hatásának egyedülállóan hatékony indikátorai is, hiszen segítségükkel olyan információk birtokába jutunk, amelyek más módszerekkel nem közelíthetők meg.
A lepkék közül – Európa negyedik legnagyobb rovarrendje – a jelenleg szakmai körökben elfogadott fajjegyzék szerint Európában 8470 lepkefaj él 35 ország, illetve sziget területén.
Csak Magyarországról közel 3500 fajt mutattak ki! Fajgazdagságuknak köszönhetően egy adott területen élő lepkék listája rendkívül plasztikusan írja le az adott terület természeti körülményeit, és ez visszafele is igaz, az abiotikus tényezők megváltozására a lepkék érzékenyen reagálnak, ezáltal jó indikátornak bizonyulnak a klímaváltozás szempontjából.

A farkasalmalepke (Zerynthia polyxena) foltszerű elterjedése tápnövénye, a farkasalma (Aristolochia clematitis) elterjedését követi (fotó: Petrányi Gergely)
A vadon élő fajok regionális elterjedésében és lokális abundanciájában jól felismerhető változások figyelhetők meg, melyek a XX. századi melegedő klimatikus trendnek tulajdoníthatók. A vizsgált lepkefajok többségének elterjedési tartományában szisztematikus északi irányú eltolódás volt megfigyelhető a XX. század folyamán. 35 faj esetében figyelték meg , hogy elterjedési területük 35-240 km-rel északabbra tolódott.

Az araszolók tökéletesen alkalmazkodtak a sziklás élőhelyekhez és ragaszkodnak is hozzájuk – változó sziklaaraszoló (Charissa variegata) (fotó: Petrányi Gergely)
A fajok élőhelyében a XX. sz. folyamán világszerte szisztematikus változás következett be, és ezek a változások összhangban voltak a globális felmelegedési szcenáriók előrejelzéseivel. Így, noha a kutatók nem tudtak kontrollált kísérleteket véghezvinni a klímaváltozásnak a fajok elterjedésére gyakorolt hatását illetőleg, a hasonló mintázatú mozgások ismétlődése az összes tanulmányban kellően erős bizonyítékát adja annak, hogy a klímaváltozás jelenleg is jelentős hatással van a természetes populációkra.

A dolomit fehérlepke (Pieris ergane) hernyója, az érintetlen dolomit sziklagyepeink jelzőfaja (fotó: Petrányi Gergely)

A halálfejes szender (Acherontia atropos) Afrikában telel, nyáron onnan vándorol hazánkba (fotó: Petrányi Gergely)

A homoki zöldbagoly (Actebia praecox) sztyeppfaunánk eltűnő fajainak egyike, utolsó példányait az 1970-es évek végén látták Magyarországon (fotó: Petrányi Gergely)

A füstös ősziaraszoló (Lignyoptera fumidaria) sztyeppreliktum, elterjedési területének nyugati határát hazánkban éri el (fotó: Petrányi Gergely)
Hasonló vizsgálatokat végeztek különböző gerinces csoportok áreatérképei alapján, oly módon, hogy értékelték a fajoknak egy pontban az áreahelyzetét (központi helyzet, peremhelyzet, áreán kívüli helyzet stb.). majd ez alapján elemezték az adott földrajzi analógiával jellemzett klímaváltozás által nem érintett, kárt szenvedett és élőnyben részesülő fajok arányát. (Bartholy et al 2012)
-
Taxa
Nem érintett
Veszteség
Új fajok
Lepke
55-81%
19-45%
19-36%
Emlős
54-21%
36-48%
10-31%
Madár
78-23%
15-59%
8-18%
Hüllő
51-13%
18-21%
28-69%
Kétéltű
75-50%
17-25%
8-25%
A hőmérséklet emelkedése és a csapadék mennyiségének egyidejű csökkenése kihat a fajok elterjedésére, befolyásolja az area határokat közvetlenül is, de a különböző fajok interakcióin keresztül is. A változások a különböző fajokra természetesen nem egyformán hatnak. Például, a mérsékeltövben a melegedés növelheti a fajok elterjedési területének méretét, hiszen a faj észak felé terjedhet, ugyanakkor a száraz, szubtrópusi területeken a csapadék csökkenése kedvezőtlenül hat, szűkíti az itteni fajok élőhelyeinek méretét.
Egymástól független kutatási eredmények azt mutatták, hogy több mint ezer vizsgált növény-, illetve állatfaj 10 év alatt átlagosan 6,1 kilométerrel mozdult el a hűvösebb területek felé. Britannia déli részén 59 madárfaj elterjedési határa átlagosan 19 km-rel tolódott el észak felé 20 év alatt. Egyes északi fajok elterjedésének északi és déli határvonala egyaránt északra tolódott, vagyis a teljes areájuk magasabb szélességre csúszott. A déli elterjedésűeknél az ellentétes tendencia volt megfigyelhető.
A madárfajok elterjedésében – feltehetően az éghajlat melegedésének hatására – bekövetkező változásokat már a múlt század elején is észleltek. A mediterraneumból ekkor indult észak felé pl. a csicsörke, amely állandó madárból – azóta Skandináviát is elérve – vonuló madár lett. A Magyarországon a ’30-as években megjelenő balkáni gerle vagy balkáni fakopáncs az ’50-60-as évekre már általánosan elterjedt, gyakori faj lett. Az előbbi azóta elérte Izlandot és Észak Afrikát is, utóbbi alig haladt túl a Kárpátokon. Mindkét betelepülő csökkentette a honos rokon fajok – a vadgerle és a nagy fakopáncs – egyedszámát, bizonyos élőhelyekről ki is szorította azokat.
Az utóbbi két-három évtizedben rohamosan nőtt a hazánkban megfigyelt új madárfajok száma. Ezek a fajok különböző állatföldrajzi területekről származnak, közöttük számos déli-délkeleti elterjedésű faj is van, amelyek megjelenése – egy-két kivételtől eltekintve – összefüggésbe hozható a klíma változásával. Az Európa déli részén növekvő egyedszámú, terjeszkedő berki poszáta első példányát 1996-ban észlelték Fenékpusztán, és 2003-tól néhány pár már költ a Kis-Balatonon.
Egyre gyakoribb a citrombillegető előfordulása is. A faj – döntő többségében hím – példányai 1989-től egyre gyakrabban kerülnek szem elé tavasszal. Kelet felől terjed, és Németországban már költ, így előbb-utóbb hazánkban is előkerül fészke. 1992-től jellemző a halászsirály, főleg ivaréretlen egyedeinek átnyaralása az ország keleti részén.
Sok esetben arról van szó, hogy a tavaszi vonulás során egyes példányok túlrepülik a fajra jellemző fészkelési területet, és attól északabbra próbálnak revírt foglalni. Ebben az esetben a madarak többsége, esetleg minden példány hím. Minél közelebb van a faj fészkelő területe, annál nagyobb az esély, hogy az ilyen hiba előfordul. Ily módon az észak felé terjeszkedő fajok esetén az első adatok általában az ilyen egyedektől származnak. Ilyen fajok pl. a sivatagi lile, pettyes kakukk, halvány sarlósfecske, vörhenyes fecske, pusztai hantmadár, kék kövirigó, bajszos poszáta, kucsmás poszáta és a kucsmás sármány.

Kucsmás poszáta ( Sylvia melanocephala) (Fotó: Csörgő T.)
A madarak vonulásuk során kihasználják a kedvező irányú szeleket, következésképpen egy, a szokottnál erősebb front el is sodorhat egy-egy madarat. Ilyen nagyon erős szél szállíthatta a Kárpát-medencébe az afrikai elterjedésű zátony kócsagot, tüskés bíbicet, de déli melegfront hozhatja a havasi sarlósfecskéket és a vékonycsőrű sirályokat is.
A klímaváltozásnak az emlősfaunára gyakorolt hatásairól jóval kevesebbet tudunk, mert hazai őshonos emlősfajaink többsége rejtett életmódú kisemlős. A médiában azonban időnként feltűnnek az árvizek kapcsán bajba jutott, vagy éppen az elől menekülő nagyvadak képei. Az aranysakál (Canis aureus) Vörös Könyves ragadozó fajunk, amely a XX. század utolsó évtizedében újra megtelepedett hazánkban. Az area expanziójának hátterében több más tényező mellett a klímaváltozás is szerepet játszhat.

Aranysakál (Canis aureus) (forrás: picasaweb)
A klímaváltozás hatással lehet továbbá a téli álmot alvó állatok (pl. a közkedvelt sün) aktivitására, telelési sikerességére is.

Sün (Erinaceus concolor) (forrás: http://gallery.photo.net)
Az extrém időjárási események és a táplálékhálózatok:
Az üvegházhatást fokozó antropogén hatásnak köszönhető tényezők, mint a talajnedvesség, a földfelület albedója jelentősen képesek befolyásolni az extrém időjárási események előfordulási gyakoriságát és mértékét, az elkövetkezendő időben egyre több extrém meleg nap, illetve csapadék várható.A klimatikus extremitásokat két csoportba szokták sorolni (Easterling et al. 2000):
- Évente rendszerint megtörténnek: Nagyon alacsony/magas napi hőmérséklet, sok napi/havi esőzés.
- Egy évben nem feltétlen fordulnak elő. Lehetnek például: aszály, árvizek, hurrikánok.
Az extrém klímaesemények szerepét szemléltetik Thibault és munkatársainak eredményei (2007). 1977 óta monitorozzák a rágcsálókat a vizsgálati területükön. Két jelentős esemény írtak le:
- 1983 : trópusi vihar 129 mm csapadékkal 6 nap alatt.
- 1999: 30 mm eső 2 óra alatt.
A vizsgálat kezdete óta változik a tájkép: több a téli csapadék, emiatt a növényzet borítottsága is megsokszorozódott. Ennek oka, hogy az El Niño jelenség hatására a télen hulló csapadék mennyisége nőtt, így a fűfélék számára kedvezőbb volt a terület. A nagyobb testű fajok idővel kiszorultak volna a bozótosabb területekre. Az 1999-es árvíz előtt a Chaetodipus baileyi faj egyedszáma exponenciálisan nőni kezdett, míg a két nagyobb tesű faj ( Dipodomys spp. Lassan csökkent). Az árvizek után nagy volt a mortalitás a két Dipodomys faj esetén (90%<), a két Chaetodipus faj esetén nem volt mérhető mortalitás, az árvizet követő hónapokban pedig jelentős növekedésbe kezdett.
Ez annak köszönhető, hogy a kengurupatkányok (Dipodomys spp.) rossz mászók és úszók. Az 1983-as hosszú esőzés másként hatott a fajokra: a napokig tartó esőzés eláztatta a talajt, ami D. spectabilis egyedeket jobban érintette (a mélyre ásott magok nem tudtak kiszáradni, megromlottak). A D. merriami ezzel szemben magasabban raktározza a magokat, amelyek így kiszáradtak: nem érintette annyira az esőzés (a D. spectabilis 1994-re eltűnt a területről.). Az esőzések ezen felül megváltoztatták a ragadozó populáció összetételét is. Olyan, addig specialista faj terjedt el
A legfőbb tanulsága az eseményeknek, hogy egy rövid idejű zavarás képes megváltoztatni egy terület életközösségeinek stabil berendezkedését, mivel több versengő faj másként képes reagálni a zavarásra.
A klímaváltozással járó, nem várt időjárás változások akár indirekt módon befolyásolhatják a predátor – préda kapcsolatokat is. A Chile félszáraz vidékein élő egérfaj (Phyllotis darwini) populációjánál is tapasztalható események szolgálhatnak például a jelenségre: Amikor az El Niño egymás utáni években is jelentkezik, a sok csapadék hatására az állatok számára elérhető táplálék. A sok táplálék a tenyészidőszakban pozitív hatással van az egerek egyedszámára, amíg az egerek egyedsűrűsége nem válik korlátozó tényezővé. Tenyészidőn kívül viszont a predátorok egyedszáma befolyásolta az egerek számát: legerősebben a gyöngybagoly (Tyto alba) (specialista) és a Culpeo pamparóka (Pseudalopex culpaeus) (generalista). Egy földibagoly faj (Speotyto cunicularia) csak akkor eszi ezeket az egereket, ha már sok bagoly van a területen és az általuk kedvelt prédák már nem állnak kellő mennyiségben a rendelkezésre (Lima et al 2002.). Ez rendszerint az El Niño időszakban be is következik.
Mint láthattuk, a klímaváltozás jelentős átalakulásokat okozhat szinte minden létező életközösségben. A probléma legfőbb oka mai napig az üvegház hatású gázok megemelkedett koncentrációja. Ez számos visszacsatolási hurok következtében jelentős átalakulást okoz Földünk klímarendszerében, megemelkedett hőmérséklettel, gyakrabban előforduló klimatikus extremitásokkal. Ezekre a változásokra próbál reagálni minden létező élőlény, kezdetben toleranciával, majd fenológiai, elterjedésbeli változásokkal. Nem minden élőlény képes megfelelő módon alkalmazkodni, ezáltal a rendszerek sok esetben érzékennyé válnak a további behatásra. Egy- egy életközösségben történt összeomlás befolyással lehet az időjárás megváltoztatására, amely tovább erősítheti a folyamatot.
Irodalomjegyzék
Asner, G. P., A. R. Townsend, B. H. Braswell: Satellite observation of El Niño effects on Amazon forest phenology and productivity, Geophys. Res. Lett., 27, 981-984, 2000.
Both, Christiaan, Sandra Bouwhuis, C. M. Lessells & Marcel E. Visser (2006): Climate change and population declines in a long-distance migratory bird - Nature 441, doi:10.1038/nature04539
Brown, P. (1998): Forest fires: setting the world ablaze - The Guardian [London], 20 March.
Bunn, A.G., S. J. Goetz, J. S. Kimball, K. Zhang (2007): Northern High-Latitude Ecosystems Respond to Climate Change - Eos, Vol. 88, No. 34
Crick Humphrey Q. P (2004): The impact of climate change on birds - Ibis (2004), 146 (Suppl.1), 48– 56
Eaton, J.G., Scheller, R.M. (1996): Effects of climate warming on fish thermal habitat in streams of the United States. – Limnol. Oceanogr. (41): 1109–1115.
Easterling, David R; Gerald A Meehl; Camille Parmesan; Stanley A Changnon;Thomas R. Karl; Linda O. Mearns (2000): Climate extremes: Observations, modeling, and impacts - Science 289, 5487
Fearnside, P. M. (2000): Global warming and tropical land-use change: greenhouse gas emissions from biomass burning, decomposition and soils in forest conversion, shifting cultivation and secondary vegetation - Climatic Change 46:115–158.
Harnos, Zs., Gaál, M., Hufnagel, L. (szerk) (2008): Klímaváltozásról mindenkinek – Budapesti Corvinus Egyetem, Budapest. (1-197 oldal) ISBN 978-963-503-384-3
Harrington R., Stork N. E. (eds) (1995): Insects in a Changing Environment. 17th Symposium of the Royal Entomological Society. London: Academic Press.
Houghton, R. A. (1991): Tropical deforestation and atmospheric carbon dioxide - Climatic Change 19:99–118.
Hufnagel L, Sipkay Cs (szerk) (2012): A klímaváltozás hatása ökológiai folyamatokra és közösségekre – Budapesti Corvinus Egyetem, Budapest (1-530 oldal) ISBN 978-963-503-511-3
Inouye D.W., Barr, B., Armitage, K.B. & Inouye, B.D. (2000):Climate change is affecting altitudinal migrants and hibernating species - PNAS 97: 1630–1633.
Kelly, Anne E. & Michael L. Goulden (2008): Rapid shifts in plant distribution with recent climate change - PNAS vol. 105 ͉ no. 33 ͉ 11823–11826
Laurance William F. and G. Bruce Williamson (2001): Positive Feedbacks among Forest Fragmentation, Drought, and Climate Change in the Amazon - Conservation Biology, Pages 1529–1535 Volume 15
Lean, J., and D. A. Warrilow (1989): Simulation of the regional climatic impact of Amazon deforestation - Nature 342:411-413.
Lima Mauricio, Nils Chr. Stenseth and Fabian M. Jaksic (2002): Population dynamics of a South American rodent: seasonal structure interacting with climate, density dependence and predator effects - Proc. R. Soc. Lond. B 269, 2579-2586 doi: 10.1098/rspb.2002.2142
Parmesan C., Yohe G. (2003): A globally coherent fingerprint of climate change impacts across natural systems. Nature, 421:37-43.
Sanz, J.J. 2002. Climate change and breeding parameters of great and blue tits throughout the western Palearctic. Global Change Biol. 8: 409 – 422.
Shukla, J., C. A. Nobre, and P. Sellers (1990): Amazon deforestation and climate change - Science 247:1322–1325.
Thibault, Katherine M. & James H. Brown (2007): Impact of an extreme climatic event on community assembly - PNAS vol. 105.͉no 9.
Thuiller Wilfried, Sandra Lavorel, Martin T. Sykes and Miguel B. Araújo (2005): Using niche-based modelling to assess the impact of climate change on tree functional diversity in Europe - Diversity and Distributions, (Diversity Distrib.) 12, 49–60
Thuiller Wilfried, Sandra Lavorel, Miguel B. Araujo, Martin T. Sykes, and I. Colin Prentice: Climate change threats to plant diversity in Europe - PNAS, doi10.1073pnas.0409902102
Végvári Zsolt, Veronika Bókony, Zoltán Barta and Gábor Kovács (2009) : Life history predicts advancement of avian spring migration in response to climate change - Global Change Biology, doi: 10.1111/j.1365-2486.2009.01876.x
Visser Marcel E. & Christiaan Both (2005): Proc. R. Soc. B 272, 2561–2569 doi:10.1098/rspb.2005.3356
Voigt, Winfried, Jörg Perner, Andrew J. Davis, Till Eggers. Jens Schumacher, Rudolf Bährmann, Bärbel Fabiani, Wolfgang Heinrich, Günter Köhler, Dorit Lichter,Rolf Marstaller, AND Friedrich W. Sander (2003) : Trophic levels are differentially sensitive to climate - Ecology, 84(9), pp. 2444–2453
![]()
![]()

Hírek/News
Sajtóközlemény
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
BővebbenSikeres pályázat
A projekt célja magyar és angol nyelvű digitális tananyagok fejlesztése a Budapesti Corvinus Egyetem Kertészettudományi Karának hét tanszékén. Az összesen 14 tananyag (hét magyar, hét angol) a kertészmérnök Msc szak és a multiple degree képzés keretében kerül felhasználásra. A digitális tartalmak az Egyetem e-learning keretrendszerével kompatibilis formában készülnek el.
A tananyagok az Új Széchenyi Terv Társadalmi Megújulás Operatív Program támogatásával készülnek.
TÁMOP-4.1.2.A/1-11/1-2011-0028
Félidő
A pályázat felidejére elkészültek a lektorált tananyagok, amelyek feltöltése folyamatban van.
![]() TÁMOP-4.1.2.A/1-11/1-2011-0028
TÁMOP-4.1.2.A/1-11/1-2011-0028